
Introduction
Over the past decades, the number of opportunistic fungal diseases has increased, taking advantage of debilitated physiological conditions or of local or systemic immune malfunction [1, 2]. Fungal infections have a particular incidence in individuals above 65 years that also display more frequently signs of immunosenescence [3]. Furthermore, inflammation increases with age [4] and can shift the balance between commensalism and infection towards the latter [5], making aged individuals more susceptible to opportunistic infections.
Candida albicans is often described as commensal, being part of the human microbiota. However, when the immune status and/or the host microbiota are altered, commensalism evolves toward extensive colonization of mucosal surfaces and disease. In these cases, gut is a main gateway for C. albicans colonization [6]. The imbalance between pro- and anti-inflammatory signals, responsible for keeping the host-fungus equilibrium, can result in severe host damage, such as chronic inflammation, but also renders fungal cells more aggressive [7]. Adenosine, through the recruitment of adenosine receptors, in particular A2A receptors (A2AR) is a main signal decreasing gut inflammation [8, 9]. This major controller of the reactivity of the immune-inflammatory system [10] undergoes a decreased capacity to dampen inflammation in some organs upon ageing [11], but it is unknown if A2AR density is modified in the gut of aged individuals.
Our objective was to understand, using a murine model, the relation between host age and susceptibility to over-colonization or infection by C. albicans, and the resulting inflammation of the gut. We also aimed to explore and correlate the localization and density of adenosine A2AR in the gastrointestinal tract with age and C. albicans over-colonization. The in vivo results indicate that ageing favors the over-colonization of the intestinal lumen and that this bolsters inflammatory signs. Moreover, aged mice display a lower A2AR density, which remains unchanged upon C. albicans infection, in contrast to young or adult mice.
Results
C. albicans load in mice stools and target organs
Since mice are not normally colonized by C. albicans, we made use of an established protocol for gastrointestinal (GI) colonization [12, 13] (Figure 1); this requires an initial elimination of the existent microbiota with antibiotics followed by infection of the mice by oral intake of a yeast suspension in the drinking water. To exclude the possibility of microbiota transference between different animals [14, 15, 16], mice were housed individually. We confirmed C. albicans identity in stools and tissues using a PCR analysis and correspondent ITS-5.8S sequencing (data not shown). In control animals (without C. albicans administration), no yeast cells were found in stools (data not shown). Using this GI infection model, none of the mice from the different groups suffered significant weight alterations (data not shown) and no mice died during the experiments, in line with previous studies [14].
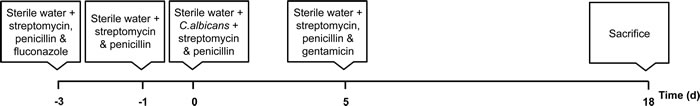
Figure 1: Schematic timeline of the gastrointestinal murine model of infection. Young, adult and aged mice were first maintained with antibiotics in the drinking water to eliminate the gut microbiota (days -3 to 0); this allows an oral infection with C. albicans (days 0 to 5) to trigger an effective colonization of the GI tract (days 5-18). Mice were sacrificed at day18 for collection of tissues for analysis.
To check GI tract colonization, the fungal load of stools was monitored every two days; as expected, at day 0, fungal cells were absent from the mice commensal flora (Figure 2A). Along the experiment, following the infection with C. albicans, the levels of yeasts elimination in adult mice remained constant, beginning to be detected at day 1, with values between 3x106 and 7x106 CFU/g of stool (Figure 2A). During the initial period of C. albicans infection per os, the stool of young and aged mice displayed similar values of fungal load, although with some point differences. In the last day of C. albicans administration (day 5), the number of yeast cells excreted by young and aged mice was significantly higher than in adult animals (~1x107 CFU/g stool, p < 0.001 and p < 0.05, respectively, vs. adult mice; Figure 2A). Between days 7 and 9, the number of viable yeast cells eliminated in the stools of young and aged mice became similar to the profile found in adults until the end of the study, except that in the last time point (day 11) where aged mice had a higher number of yeast cells in the stools when compared with the other age groups (~2x107 CFU/g stool, p < 0.001; Figure 2A).
At the end of the study, the animals were sacrificed and the fungal content was investigated in kidneys, livers, stomachs, small and large intestines. No yeast colonization was found in either kidneys or livers from any group (Figure 2B). C. albicans cells were found in all the gastrointestinal tract tissues, stomach, small intestine and large intestine from infected mice of the three age groups. The results for fungal content had high variability (values ranging from ~8x104 to ~1x106 CFU/g of tissue), and the samples taken from the large intestine of the three age groups displayed larger fungal loads than the other GI tissues (~1x106 CFU/g tissue; Figure 2B). However, when comparing between the three groups, aged mice tend to present a lower colonization level in the stomach and intestine, although not statistical significant compared to adult mice (Figure 2B). The histological analysis revealed that, at day 11, the colonization of the stomach mucosa was more evident than in the intestine (small and large). Interestingly, C. albicans yeast-to-hyphae transition in the stomach was more exuberant in aged mice than in young and adult mice (Figure 2C).
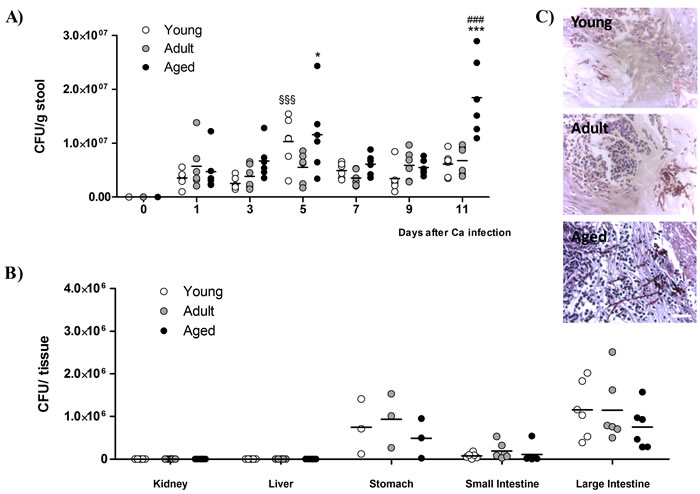
Figure 2: Yeast load in mice stools and target organs. A. Stool fungal burdens of young, adult and aged mice before (day 0) and in consecutive days after C. albicans oral infection. Stools were collected every two days until the end of the experiment, from mice housed individually. B. Viable C. albicans counts in different tissues of young, adult and aged mice at the end of the experimental procedure (day 18). Dots represent data from individual mice and horizontal lines represent the mean of each group of mice; * p < 0.05, *** p < 0.001, aged vs. adult mice; ### p < 0.001, aged vs. young mice and §§§p < 0.001, adult vs. young mice. C. C. albicans hyphal forms were found in stomachs from young, adult and aged mice 18 days after infection with C. albicans. Scale bar: 5 µm.
Inflammation processes in the GI tract
The analysis of the histological sections of the different GI tract tissues from control mice of the three age groups did not reveal any significant age-dependent alterations (Figures 3 and 4). According to the inflammatory criteria used to calculate a score index, we found a score of 0 (Table 1), with the absence of alterations in the tissues sections from control mice. By contrast, Candida infection induced a severe degree of disruption of the villi, haemorrhagic damage, and increase of inflammatory cells infiltration of the gastric mucosa (Figures 3; “Infected” panels). Infected aged animals had large inflammatory infiltrates (Figure 3A), and infected adult animals displayed the presence of lymphoid tissue (Figure 3B). In the small intestine, there was a mild disruption of villi and inflammatory infiltrate in the aged infected group, which did not reach significance compared to control mice (Figure 3 and Table 1). The large intestine from aged infected mice displayed mild epithelial cell loss and moderate haemorrhage (Figure 3 and Table 1). The alterations found in small and large intestines of infected mice from the different age groups revealed index scores of 0.5 to 2 (Table 1). When looking at stomach sections of young, adult and aged mice, the presence of C. albicans filamentous forms induced more severe alterations of villi, haemorrhage and inflammatory infiltrates (Table 1, score from 2 to 3), confirmed by the inflammatory processes illustrated in Figure 3. We did not observe differences between the groups with respect to the presence of glycogen or mucopolysaccharides in foveolar (stomach) and goblet cells (intestine) or presence of mucus when tissues were stained with PAS (Figure 4). Thus, both ageing and infection increased signs of inflammation in the GI tissues of the mice (Figures 3 and Table 1).
Table 1: Histological evaluation scores from mice gut tissues.
Control |
Infected |
||||
Vilos. |
Hem. Infilt. |
Vilos. |
Hem. Infilt. |
||
Stomach |
Aged |
0 (0-0) |
0 (0-0) |
2 |
3 |
Adult |
0 (0-0) |
0 (0-0) |
3 |
3 |
|
Young |
0 (0-0) |
0 (0-0) |
3 |
3 |
|
Small Intestine |
Aged |
0 (0-0) |
0 (0-0) |
1 (0-1) |
1.5 (1-2) |
Adult |
0 (0-0) |
0 (0-0) |
0.5 (0-1) |
1 (0-1) |
|
Young |
0 (0-0) |
0 (0-0) |
0.5 (0-1) |
0.5 (0-1) |
|
Large Intestine |
Aged |
0 (0-0) |
0 (0-0) |
1 (0-1) |
2 (0-2)* |
Adult |
0 (0-0) |
0 (0-0) |
0 (0-1) |
1 (1-2) |
|
Young |
0 (0-0) |
0 (0-0) |
1 (0-1) |
0 (0-0)# |
|
Histological sections of stomachs, small and large intestines of control and infected mice groups (aged, adult and young) were analysed regarding villi (Vilos.), haemorrhage (Hem.) and inflammatory infiltrates (Infilt.).
Scores represent absence (0) to mild (1) and severe (3) alterations. *, p < 0.05 - adult infected vs. aged infected; #, p < 0.05 – aged infected vs. young infected.
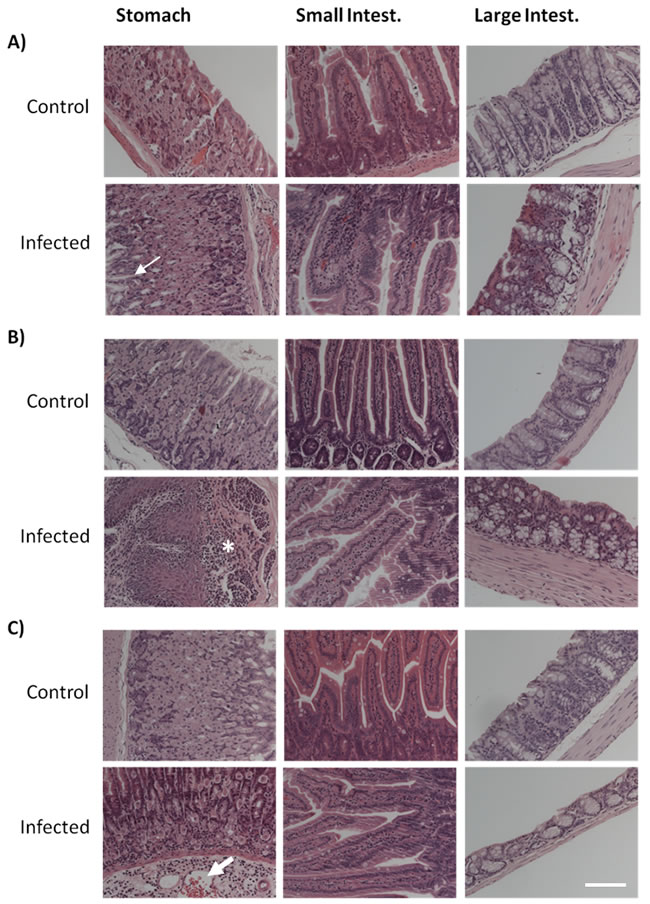
Figure 3: Histological analysis of haematoxylin-eosin (HE) stained tissues. Stomach, small and large intestine sections from both control and C. albicans infected mice from A. aged, B. adult and C. young groups. Images from infected mice illustrate C. albicans infection capacity to induce severe degree of villi disruption, hemorrhagic damage and inflammatory cells infiltration on gastric mucosa, particularly evident in aged animals. Photographs point to the yeast-induced epithelial cell loss (thin arrow), inflammatory infiltrate (asterisk), and hemorrhage (large arrow). Images were taken using a Zeiss Axio Imager Z2 Microscope; scale bar: 50 µm.
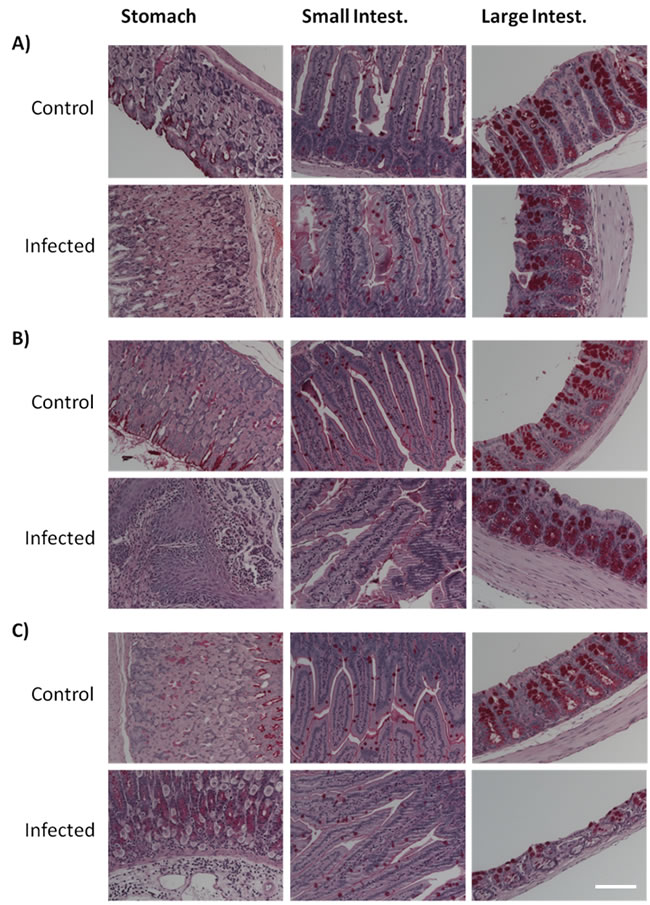
Figure 4: Histological analysis of periodic acid Schiff (PAS) stained tissues. Stomach, small and large intestine sections from both control and C. albicans infected mice from A. aged, B. adult and C. young groups. No differences in glycogen and presence of mucopolysaccharides were observed between groups. Images were taken using a Zeiss Axio Imager Z2 Microscope; scale bar: 50 µm.
Mapping A2A receptors (A2AR) in the gut of infected mice
Since infection situations trigger an up-regulation of A2AR, which feedback to curtail inflammation [10], we investigated if there was a correlation between A2AR density, inflammation, tissue damage and age. Adult and young animals displayed a larger A2AR immunoreactivity in GI tissues sections upon infection with C. albicans compared to controls (Figure 5B, 5C). This A2AR staining was particularly evident in the gastric epithelium layers of both adult and young mice, and also appeared with high intensity in the muscularis mucosa of stomach and small and large intestines from adult and young infected mice (Figure 5B, 5C, 5D). Notorious differences were observed when analyzing GI tissues from aged mice: Figure 5 shows that A2AR immunofluorescence was lower in GI tissues of aged mice compared to young and adult mice. Furthermore, the dynamic plasticity of A2AR density upon infection was lost in aged mice: C. albicans infection failed to increase A2AR staining in aged rats compared to non-infected aged rats (Figure 5A), in contrast to what was observed in young and adult mice (Figure 5B, 5C).
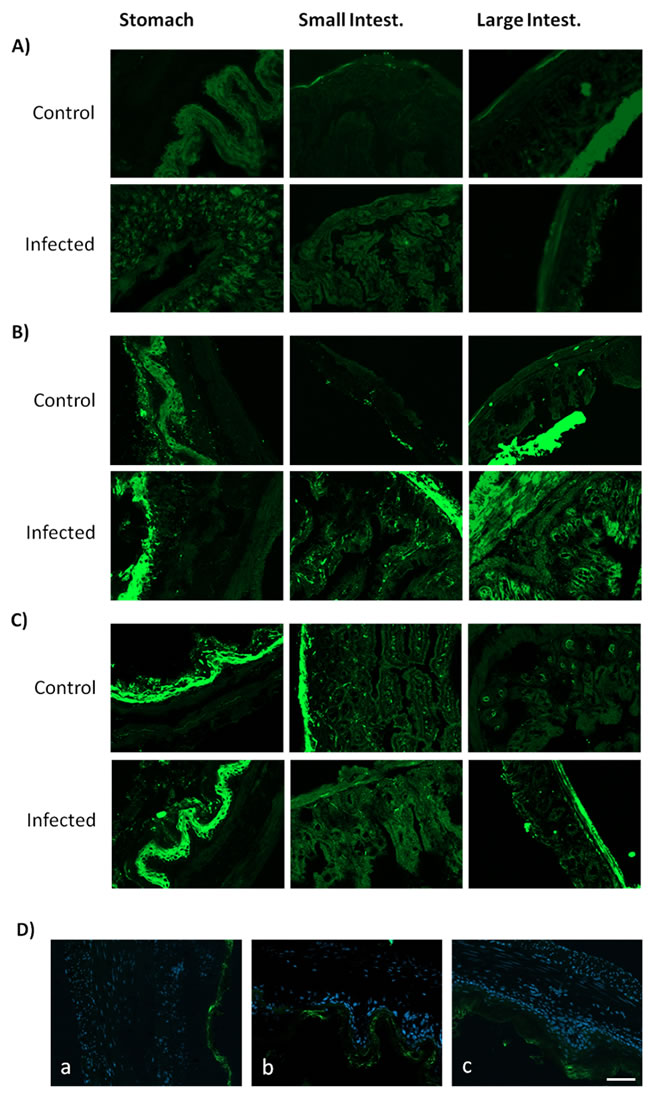
Figure 5: Adenosine A2A receptors localization on mice GI tract tissues. Representative images of stomach, small and large intestine sections immunostained with an antibody against A2AR (green) of A. aged, B. adult and C. young mice. In adults and young mice, increased A2AR density during C. albicans infection was observed. In contrast, aged mice have lower levels and alterations of A2AR immunolabeling between conditions. D. Infected stomachs of a) aged, b) adult and c) young mice showing an increased A2AR density on epithelium and muscularis mucosa layers (cell nuclei stained blue) of young and adult compared to aged mice. Images were obtained using a Zeiss Axio Imager Z2 Microscope with ApoTome2 structured illumination acquisition system; scale bar: 10 µm.
Discussion
This study tested the susceptibility to C. albicans over-growth or over-colonization of the gastrointestinal (GI) tract of mice with different ages. We aimed to correlate the over-colonization by C. albicans and subsequent inflammation in the GI tissues of young, adults and aged mice, with the modifications of adenosine A2A receptors (A2AR), a major stop signal of inflammation. We used a well established and sustained murine GI infection model [14, 15, 16] based on the elimination of mice microbiota, which allowed an effective C. albicans GI colonization, without mice death owed to infection, in all age groups. C. albicans infection/over-colonization of elderly mice stomach lead to a more exuberant yeast-to-hypha transition with higher inflammation when compared to the other age groups. We also found that the overgrowth of C. albicans in the intestinal lumen of aged mice induced an inflammation of the mucosal tissues, which was independent on the invasion of intestinal tissues by yeast cells. The pattern of A2AR distribution in the GI tract showed that aged mice have a lower A2AR density in the GI tract tissues. Furthermore, A2AR density did not increase in response to C. albicans over-growth/infection, in contrast to the increased A2AR density observed in young and adult mice upon yeast infection.
The major conclusion of our study is that aged animals were more prone to yeast over-growth in the gut, which displayed a heighted inflammatory profile. First, we observed that C. albicans over-colonization of the gastric compartment (during the first 11 days of infection) in aged mice proceeds with a more exuberant morphological switch to the hyphal form, when compared with young and adult mice. Concomitantly, higher inflammation is observed in the gastric tissues of elderly mice. The importance of C. albicans morphologic switch for infection and dissemination in the gut is well recognized and has been lately highlighted [17, 18]. We observed a higher morphological switch and over-colonization of the stomach tissues than that observed in the intestinal segments, where the fungus was predominantly located in the intestinal lumen. This was attested by the elimination of high levels of viable C. albicans cells in the stools and low levels of intestinal tissue colonization. This novel aspect of the gut infection/colonization by C. albicans was also observed by Brown and co-workers [14, 15], and strongly suggests that the intestinal inflammation results from an overgrowth of yeast cells in the lumen rather than an invasion of the intestinal tissue. This was particularly evident in the aged mice.
It has been described that inflammation can facilitate translocation phenomena (e.g., [19, 20]), and that the colonization of the gut by yeasts is associated with translocation and subsequent hematogenous dissemination in immunosuppressed mice [13, 20]. In the present work we did not observe an invasive infection of the intestinal tissues by C. albicans. This prompts the conclusion that the inflammation of the intestine, especially in aged mice, can be triggered by the recognition of cell wall components of C. albicans or by extracellular secretion of products such as farnesol [21] or small peptides [22].
In contrast with previous findings regarding systemic C. albicans infection [22], we have found low yeast levels in the kidneys and livers of all animals, using an infection model via the drinking water. Those previous studies of invasive infections [22] report a substantial decrease of the colonization levels in kidneys and livers throughout the entire period of infection, with residual values over the end of the experiment. This suggests an efficient immunity response [22]. Since we only harvested the tissues at the end of the experiment, the low fungal burden quantified corresponds to a post-eradication period. Nevertheless, the low colonization rates that we now observed, were also previously reported by others [15, 23, 24].
The second major finding of our study is the tentative association of A2AR in GI tissues with the over-colonization by C. albicans and subsequent inflammation. The involvement of purines and their receptors in the pathophysiology of inflammatory gut diseases is best heralded by combined observations describing that, while extracellular ATP mediates inflammation [25], CD73, an ecto-nucleotidase responsible for converting AMP in adenosine, increases the recovery rate from severe colonic inflammation [26]. Adenosine is recognized as an anti-inflammatory agent, contributing to ameliorate the damage induced by inflammation [10, 27]. Accordingly, A2AR are involved in inhibitory mechanisms of pro-inflammatory cytokines secretion during colitis or enteritis, and the levels of adenosine in the intestinal luminal are increased in the inflamed intestine in several pathological conditions such as inflammatory bowel disease [28]. Our study provides the first qualitative analysis of A2AR distribution in the GI tract of mice from different age groups. We observed that aged mice had a lower A2AR density than young and adult mice. Moreover, instead of increasing the density of A2AR during C. albicans infection, as observed in adults and young mice, A2AR immunolabelling showed that A2AR density was maintained or slightly decreased. Therefore, we conclude that the inability to increase A2AR density in the GI tract of infected aged mice, contributes to a higher inflammatory score, with a lower ability to overcome the deleterious effects of C. albicans over-colonization, leading to a higher tissue damage. Overall, this supports the hypothesis that increased A2AR density at the surface of GI mucosa cells is a protective mechanism against C. albicans overgrowth; this mechanism is present throughout adulthood and decreases with age. It would be interesting to study the effect of drugs such as rapamycin, simultaneously displaying anti-aging [29] and anti-fungal [30] properties, over C. albicans colonization and A2AR distribution.
We also observed that in aged mice, yeast-to-hypha transition of C. albicans was more exuberant, especially in the stomach. We propose that this may result from the lower density of A2AR because adenosine and A2AR in the stomach contribute to decrease the secretion of gastric acid [31]. The lower A2AR density in the stomach of aged mice is expected to decrease the pH, leading to a stressful condition that triggers the morphological switch of C. albicans. In fact, it was described that, under acidic conditions, C. albicans raises the pH to neutral or alkaline, resulting in yeast-to-hypha transition, considered one of its most important virulent traits [32]. The age-related decrease of A2AR density observed by us, together with the ability of C. albicans to modulate the external pH [32], might explain our finding of a low rate of yeast-to-hypha transition in young mice, increasing in adults, and displaying a maximal rate in aged mice. This novel working hypothesis is of particular interest in the context of gastrointestinal diseases, which have increased incidence in elderly patients [33], especially in those with gastric ulcers who are more colonized by C. albicans [34].
In summary, the overgrowth of C. albicans in the intestinal lumen of aged mice induced inflammation of the mucosal tissues, which was independent on the invasion of intestinal tissues by yeast cells. The pattern of A2AR distribution in the GI tract showed that aged mice have less A2AR in the GI tract tissues, and that A2AR density does not increases in response to C. albicans overgrowth/infection, in contrast to the increased A2AR density observed in young and adult mice upon yeast infection. This paves the way to exploit the purinergic system, in particular the adenosine A2A receptor, associated with beneficial effects under inflammatory conditions, to control over-colonization/infectious process of C. albicans in the gut of elderly. The impact of these findings is that the lack/reduced number of A2AR likely contributes to increased inflammation and lower ability to decrease C. albicans (or other) gut infection and the correspondent deleterious effects in the elderly.
Materials and Methods
Strain, media and growth conditions
C. albicans YP0037 strain (Microbiology Pathogenic Yeast Collection, University of Coimbra) was isolated from a hemoculture. Yeast cells were grown overnight at 30ºC on YPD agar (0.5% yeast extract, 1% bacto-peptone, 2% agar and 2% glucose) plates, harvested by centrifugation and resuspended in PBS (phosphate buffer saline, pH 7.4). Cell density was determined using a Neubauer chamber cell.
Mice and gastrointestinal infection model
Young, adult and aged (respectively, 2-, 9- and 18-months-old) C57Bl/6JRj male mice were obtained from Janvier Labs (France) and maintained under specific pathogen-free conditions at the animal facilities of the Faculty of Medicine, University of Oporto. All procedures were according to EU guidelines (2010/63) and approved by the Directorate General of Food and Veterinary Medicine of the European Union (authorization no. 6411). Animals were individually housed in sterilized cages, supplied with sterile bedding, water and mouse chow and kept under a 12h:12h light-dark cycle, under controlled temperature (21-24ºC) and humidity (50-60%). The gastrointestinal infection model was carried out as previously described [14] with slight modifications (Figure 1). Briefly, to reduce commensal bacterial and fungal flora, mice were treated for 3 days with sterile drinking water containing 2 mg/mL streptomycin (Reig Jofré, Spain), 2000 U/mL penicillin G (Atral, Portugal) and 0.25 mg/mL fluconazole (Pfizer, Germany) and then switched for a further day to water containing the same concentrations of streptomycin and penicillin G. Then, animals were provided, during 5 days, with sterile drinking water with C. albicans (1x107 colony forming units, CFU/mL) and the same concentrations of streptomycin and penicillin G. After this period of yeasts ingestion, mice were maintained with sterile water containing 2 mg/mL streptomycin, 2000 U/mL penicillin G and 0.2 mg/mL gentamicin (Labesfal, Portugal) until the end of the experiment. Two additional groups of mice were maintained without C. albicans administration, one of the groups kept under the same antibiotic protocol while the other group was not exposed to antibiotics. To confirm the depletion of the endogenous microflora, stools were collected before beginning C. albicans administration.
To monitor C. albicans colonization the stools were collected from individual mice every two days. Stools were homogenized in 1 mL PBS and serial dilutions were made in order to obtain 30 to 50 CFU in each YPD agar plate. Plates were incubated for 2 days at 30 ºC and the yeast load was quantified by counting the number of colonies corresponding to viable yeast cells and expressed as CFU per g of stools. Mice were sacrificed 19 days after C. albicans exposure and kidneys, livers, stomachs, small intestines and large intestines were harvested and weighted. Equivalent sets of tissues were divided into three groups, (i) fixed in formalin solution for subsequent histology analysis; (ii) immediately snap-frozen in isopentane and kept at -80 ºC, for further immunohistochemistry; (iii) homogenized in PBS with 0.05% Triton X-100, diluted and plated on YPD as described above, to determine the tissue content of C. albicans.
Histology
Formalized tissues were processed using standardized paraffin embedding protocols and stained with hematoxylin and eosin (HE) and periodic acid-Schiff (PAS) methods. Sections of the different organs were microscopically analyzed (Zeiss Axio Imager Z2 Microscope, with EC Plan-Neofluar, 5x/0.16 and 40x/0.75, objectives) in order to evaluate the presence of yeasts and/or inflammation. Stomach and small intestine sections were evaluated according to the criteria of Kelly [3]. Briefly, a 1-cm segment of each histological section was assessed on a scale of 0 (absence of alterations) and 1 (mild) to 3 (severe) for disruption of the villi, presence of red blood cells, and neutrophil infiltration. For large intestine, the presence and intensity of histological changes were evaluated with the use of a score index, according to Vilaseca [36], including the following criteria: presence of epithelial cell loss, presence and intensity of inflammatory infiltration and presence of blood cells.
Immunohistochemistry
Snap-frozen samples were analysed using an immunohistochemistry standard protocol against A2AR. Briefly, OCT (Tissue-Tek, Sakura) ultrathin (10 µm) sections (Cryostat CM3050S, Leica Biosystem) were obtained from stomach, small and large intestine of the different mice. Tissue sections were fixed with pre-cooled acetone (-20°C) for 10 min, washed with PBS and permeabilized with PBS and 0.25% Triton X-100 for 30 min. After PBS washing, 10% albumin bovine serum (BSA) solution in PBS was used as blocking buffer and sections’ slides were incubated in a humidified chamber at room temperature for 1 h. The rabbit anti-A2AR primary antibody (1:300; sc-13937 from Santa Cruz Biotechnology) was diluted in PBS with 1% BSA and incubated with the sections overnight at 4°C in a humidified chamber. Sections were then washed with PBS, and treated with the Alexa Fluor 488-labeled donkey anti-rabbit secondary antibody (1:2000; A21206 from Molecular Probes) diluted in PBS with 1% BSA. Sections were again incubated in a humidified chamber at room temperature for 2 h. After washing with PBS, nuclei were counterstained with DAPI (Sigma), during 5 min at room temperature, protected from light, and again rinsed with PBS. Coverslips were mounted with Vectashield HardSet mounting medium and kept at 4 ºC until visualized with fluorescence and/or confocal microscopy. Digital images were captured using a Zeiss Axio Imager Z2 Microscope (with ApoTome2 structured illumination acquisition system and a Plan-Apochromat 20x/0.8 objective), and using Zeiss ZEN 2012 (blue edition) and ImageJ software was used to analyse the images.
Statistics
Data are means ± SEM. Statistical differences were determined with one or two-way ANOVA respectively followed by a Newman-Keuls or Bonferroni post hoc test. Histological damage scores were analysed with Kruskall Wallis and Mann-Whitney tests. Statistical significance was considered at p < 0.05.
Acknowledgments
We thank Professor Maria de Fátima Carneiro and Rui M. Correia from the Department of Anatomic Pathology, University Hospital of S. João, Porto, for the collaboration in the histological procedures.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Funding
This work was supported by Fundação para a Ciência e Tecnologia (FCT) grants (PTDC/SAU-FCF/81436/2006; PTDC/SAU-MIC/115598/2009), by FEDER funds through the Operational Programme Competitiveness Factors - COMPETE and national funds by FCT -Foundation for Science and Technology under the strategic project UID / NEU / 04539 / 2013; and also by NARSAD. LR is a recipient of a PhD grant by FCT (SFRH/BD/74181/2010). IMM is supported by FCT (BPD/113285/2015), FEDER and COMPETE.
References
1. Falagas ME, Roussos N and Vardakas KZ. Relative frequency of albicans and the various non-albicans Candida spp among candidemia isolates from inpatients in various parts of the world: a systematic review. Int J Infect Dis. 2010; 14:e954-e966.
2. Pfaller MA and Diekema DJ. Epidemiology of invasive candidiasis: a persistent public health problem. Clin Microbiol Rev. 2007; 20:133-163.
3. Aw D, Silva AB and Palmer DB. Immunosenescence: emerging challenges for an ageing population. Immunology. 2007; 120:435-446.
4. Biagi E, Nylund L, Candela M, Ostan R, Bucci L, Pini E, Nikkïla J, Monti D, Satokari R, Franceschi C, Brigidi P and De Vos W. Through ageing, and beyond: gut microbiota and inflammatory status in seniors and centenarians. PLoS ONE. 2010; 5:e10667.
5. Round JL and Mazmanian SK. The gut microbiota shapes intestinal immune responses during health and disease. Nat Rev Immunol. 2009; 9:313-323.
6. d’Enfert C. Hidden killers: persitence of opportunistic fungal pathogens in the human host. Curr Opin Microbiol. 2009; 12: 358-364.
7. Romani L. Immunity to fungal infections. Nat Rev Immunol. 2011; 11:276-288.
8. Estrela AB and Abraham WR. Adenosine in the inflamed gut: a Janus faced compound. Curr Med Chem. 2011; 18:2791-2815.
9. Haskó G, Linden J, Cronstein B and Pacher P. Adenosine receptors: therapeutic aspects for inflammatory and immune diseases. Nat Rev Drug Discov. 2008; 7:759-770.
10. Sitkovsky M, Lukashev D, Apasov S, Kojima H, Koshiba M, Caldwell C, Ohta A and Thiel M. Physiological control of immune response and inflammatory tissue damage by hypoxia-inducible factors and adenosine A2A receptors. Annu Rev Immunol. 2004; 22:657-682.
11. Fredholm BB, Chen JF, Cunha RA, Svenningsson P and Vaugeoisx JM. Adenosine and brain function. Int Rev Neurobiol. 2005; 63:191-270.
12. Cole GT, Halawa AA and Anaissie EJ. The role of the gastrointestinal tract in hematogenous candidiasis: From the laboratory to the bedside. Clin Infect Dis. 1996; 22:873-888.
13. Koh AY, Köhler JR, Coggshall KT, Van Rooijen N and Pier GB. Mucosal damage and neutropenia are required for Candida albicans dissemination. PLoS Pathog. 2008; 4:e35.
14. Vautier S, Drummond RA, Redelinghuys P, Murray GI, MacCallum DM and Brown GD. Dectin-1 is not required for controlling Candida albicans colonization of the gastrointestinal tract. Infect. Immun. 2012; 80:4216-4222.
15. Vautier S, Drummond RA, Chen K, Murray GI, Kadosh D, Brown AJP, Gow NA, MacCallum DM, Kolls JK and Brown GD. Candida albicans colonization and dissemination from the murine gastrointestinal tract: the influence of morphology and Th17 immunity. Cell Microbiol. 2014; 17:445-450.
16. Mason KL, Erb Downward JR, Mason KD, Falkowski NR, Eaton KA, Kao JY, Young VB and Huffnagle GB. Candida albicans and bacterial microbiota interactions in the cecum during recolonization following broad-spectrum antibiotic therapy. Infect Immun. 2012; 80:3371-3380.
17. Gow NAR. A developmental program for Candida commensalism. Nature Genet. 2013; 45: 967-968.
18. Pande K, Chen C and Noble SM. Passage through the mammalian gut triggers a phenotypic switch that promotes Candida albicans commensalism. Nature Genet. 2013; 45:1088-1091.
19. Andrutis, KA, Riggle PJ, Kumamoto CA and Tzipori S. Intestinal lesions associated with disseminated candidiasis in an experimental animal model. J Clin Microbiol. 2000; 38:2317-2323.
20. Takahashi K, Kita E, Konishi M, Yoshimoto E, Mikasa K, Narita N and Kimura H. Translocation model of Candida albicans in DBA-2/J mice with protein calorie malnutrition mimics hematogenous candidiasis in humans. Microb Pathog. 2003; 35:179-187.
21. Ghosh S, Howe N, Volk K, Tati S, Nickerson KW and Petro TM. Candida albicans cell wall components and farnesol stimulate the expression of both inflammatory and regulatory cytokines in the murine RAW264.7 macrophage cell line. FEMS Immunol Med Microbiol. 2010; 60:63-73.
22. Murciano C, Villamón E, Yáñez A, O’Connor JE, Gozalbo D and Gil ML. Impaired immune response to Candida albicans in aged mice. J Med Microbiol. 2006; 55:1649-1656.
23. Kennedy MJ and Volz PA. Effect of various antibiotics on gastrointestinal colonization and dissemination by Candida albicans. Sabouraudia. 1985; 23:265-273.
24. Samonis G, Anaissie EJ, Rosenbaum B and Bodey GP. A model of sustained gastrointestinal colonization by Candida albicans in healthy adult mice. Infect Immun. 1990; 58:1514-1517.
25. Kurashima Y, Kiyono H and Kunisawa J. Pathophysiological role of extracellular purinergic mediators in the control of intestinal inflammation. Mediators Inflamm. 2015; 2015:427125.
26. Bynoe MS, Waickman AT, Mahamed DA, Mueller C, Mills JH and Czopik A. CD73 is critical for the resolution of murine colonic inflammation. J Biomed Biotechnol. 2012; 2012:260983.
27. Haskó G and Pacher P. A2A receptors in inflammation and injury: lessons learned from transgenic animals. J Leukoc Biol. 2008; 83:447-455.
28. Ye JH and Rajendran V. Adenosine: an immune modulator of inflammatory bowel diseases. World J Gastroenterol. 2015; 15:4491-4498.
29. Blagosklonny MV. Rejuvenating immunity: “anti-aging drug today” eight years later. Oncotarget. 2015; 6:19405-19412. doi: 10.18632/oncotarget.3740.
30. Bastidas RJ, Shertz CA, Lee SC, Heitman J and Cardenas MA. Rapamycin exerts antifungal activity in vitro and in vivo against Mucor circinelloides via FKBP12-dependent inhibition of Tor. Eukaryotic Cell. 2015; 11:270-281.
31. Yip L and Kwok YN. Role of adenosine A2A receptor in the regulation of gastric somatostatin release. J Pharmacol Exp Ther. 2004; 309:804-815.
32. Vylkova S, Carman AJ, Danhof HA, Collette JR, Zhou H and Lorenz MC. The fungal pathogen Candida albicans autoinduces hyphal morphogenesis by raising extracellular pH. MBio. 2011; 2:e00055-11.
33. Holt PR. Gastrointestinal diseases in the elderly. Curr Opin Clin Nutr Metab Care. 2003; 6:41-48.
34. Kumamoto CA. Inflammation and gastrointestinal Candida colonization. Curr Opin Microbiol. 2011; 14: 386-391.
35. Kelly CP, Pothoulakis C and LaMont JT. Clostridium difficile colitis. N Engl J Med. 1994;330, 257-262.
36. Vilaseca J, Salas A, Guarner F, Rodríguez R, Martínez M and Malagelada JR. Dietary fish oil reduces progression of chronic inflammatory lesions in a rat model of granulomatous colitis. Gut. 1990; 31:539-544.