
Introduction
After T cell antigen receptor activation, the fate of naïve T cells is determined to a large extent by the cytokine environment [1-4]. Naïve CD4+T cells differentiate into functionally distinct subsets, including IFNγ-producing TH1 cells, IL-4-producing TH2 cells, IL-17-producing TH17 cells, and induced regulatory T cells (iTreg); these cells are responsible for different types of T cell immunity and affect immune-mediated responses to disease [5-8] (Figure 1). Among these T cell subtypes, IL-9-producing TH9 cells were first described in 1994 [9] and defined in 2008 [10, 11]. Initially, TH9 cells were thought to be associated with TH2 responses and to arise from TGFβ-induced reprogramming of TH2 cells [12-14]. However, recent reports have shown that TH9 cells are involved in antimicrobial immunity [15, 16], autoimmune disease [17-19], colitis [20, 21], and even anti-tumor immunity [12, 22]. This review summarizes the emerging role of TH9 cells in immune-mediated diseases, with a special focus on several recent anti-tumor immunological studies.
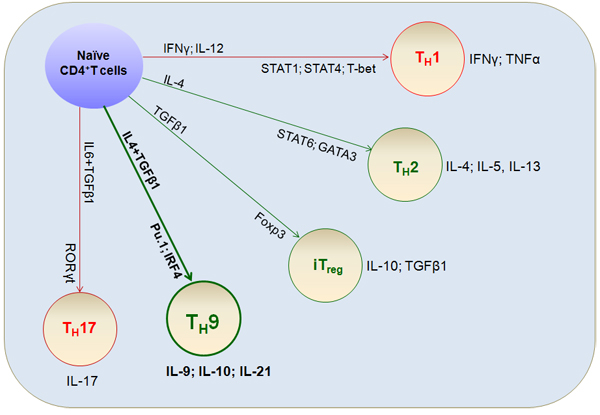
Figure 1: Differentiation of T cell lineages. Naïve CD4+T cells are activated by T cell receptor (TCR) signaling and differentiate into various T cell lineages depending on the cytokine environment. Prototypical differentiation cytokine sets, corresponding specific transcriptional factors, and functional cytokine effects that regulate TH cell fate and functions (including TH9 cells) are shown.
TH9 cell differentiation and signal transduction
TH9 cell differentiation
The cytokine milieu plays a crucial role in T cell differentiation. In addition to IL-4 and TGFβ [10, 11], IL-1 [23], IL-2 [24], IL-6 [25], IL-10 [26], IL-21 [27, 28], IL-23 [29], IL-25 [30], IL-33 [31], IFN-α/β [27], and thymic stromal lymphopoietin (TSLP) [32] promote IL-9 production in naïve T cells, while IFN-γ and IL-27 suppress IL-9 production [18]. For example, naïve CD4+T cells primed with IL-4 and TGFβ1 secrete IL-9 [9]. Compared to stimulation with TCR alone, the addition of TGFβ1 increased IL-9 production in murine CD4+T cells; IL-9 production was further increased by the addition of both TGFβ1 and IL-4, which, by itself, has scant effect [33-36]. A variety of cytokines are now known to affect Th9 cell differentiation and IL-9 production in T cells.TH9 cell development requires the integration of multiple signals, and a complex cytokine milieu is required for optimal IL-9 production; the balance of cytokine signals is therefore critical for inducing TH9 cell development and differentiation rather than the generation of other T-helper subsets.
Transcriptional control of TH9 cell differentiation
Certain transcription factors, including STAT [37], PU.1 [21, 38, 39], IRF1 [28], IRF4 [40], NF-kB [41], Bcl6 [42], and the Smad/Notch complex [43], directly interact with the Il9 gene promoter to increase IL-9 production (Figure 2). Moreover, acetylation and H3K27 trimethylation, a suppressive chromatin modification, are increased at the IL-9 promoter in TH9 cells [39, 44], resulting in barely detectable IL-9 production in these cells compared to other CD4+T cell lineages.
IL-4 and TGFβ1, and their downstream transcriptional targets, are required for TH9 cell differentiation [23, 45]. For example, IL-4-induced activation of STAT6 and the STAT6 target gene GATA3 are both required for TH9 differentiation, although GATA3 is more important for TH2 differentiation [13, 46]. Upon activation, phosphorylated STAT6 facilitates the transcription of GATA3 and IRF4 [47]. However, modest retrovirus transduction-induced expression of IRF4 and/or GATA3 did not rescue IL-9 secretion in STAT6-deficient CD4+T cells, indicating that additional factors are required for the STAT6-dependent transcriptional modulation of TH9 differentiation [46]. In addition, GATA3 transcription is activated in a STAT6-independent manner during TH9 differentiation. Notch1- and Notch2-deficient TH9 cells exhibit decreased IL-9 production; Jagged2 is able to induce IL-9 production in the presence of TGFβ1 alone in these cells, and exogenous IL-4 rescues Notch deficiency [48, 49]. The DNA-binding inhibitor Id3 inhibits IL-9 production in CD4+T cells in a GATA3-dependent manner [33]. Deletion of Id3 increases IL-9 production in CD4+T cells, indicating that Id3 also inhibits TH9 differentiation in an IL-4-GATA3-dependent manner. These data suggest that STAT6 signaling is not absolutely necessary for the induction of TH9 differentiation; Notch or Id3-mediated induction of GATA3 is sufficient.
TGFβ is also required for TH9 generation. Accordingly, the TGFβ downstream target factor SMAD is critical for TH9 cell differentiation. Binding of TGFβ to its receptor activates specific SMAD family members, and TGFβ-activated phosphor-SMAD3 directly binds to the Il9 locus, the Notch intracellular domain (NICD), and RBP-Jk (recombination signal binding protein for immunoglobulin kappa J region) [10, 43]. In addition, TGFβ1 induces transcriptional factor PU.1 expression and inhibits the expression of T-bet, a TH1-specific transcriptional factor, thereby promoting TH9 differentiation [21, 39]. PU.1 is expressed specifically in subpopulations of TH2 cells with low IL-4 expression. PU.1-deficient T cells produce less IL-9, and ectopic expression of PU.1 increases IL-9 production. Reduced PU.1 expression in human IL-9-secreting T cell cultures also reduced IL-9 production. Mechanistic studies have shown that PU.1 likely influences TH9 differentiation by interfering with GATA3 activation or by recruiting the histone acetyltransferase (HAT) proteins Gcn5 and PCAF to the Il9 locus [21, 38].
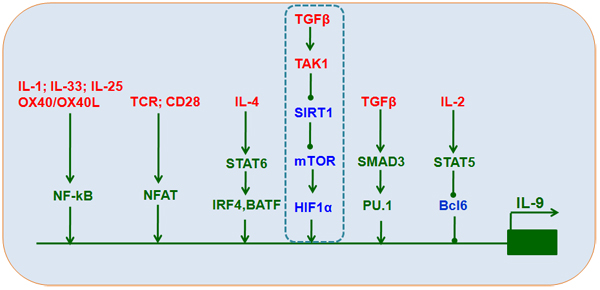
Figure 2: Transcriptional control of TH9 cell differentiation. IL-4, TGFβ1, IL-2, TCR, and other stimuli induce the expression of downstream transcriptional factors, including STAT6, IRF4, BATF, SMAD3, TAK1-SIRT1-mTOR-HIF1α, PU.1, STAT5, Bcl6, NF-kB, and NFAT, that interact with Il9 promoters to increase IL-9 expression and secretion.
The TGFβ-activated kinase TAK1 is an important mediator of Smad-independent TGFβ signaling [50] and plays a key role in directing TH9 differentiation [33]. Our recent studies confirm that TAK1 inhibition reversed SIRT1 suppression, suggesting that a Smad-independent TAK1 signal is responsible for SIRT1 suppression during TH9 differentiation. SIRT1 deficiencies induced by either conditional deletion in mouse CD4+T cells or small interfering RNA (siRNA) in mouse or human T cells increased, while ectopic SIRT1 expression inhibited, IL-9 production. Additionally, glycolytic activation through the mTOR-hypoxia-inducible factor-1α (HIF1α) pathway was required for TH9 cell differentiation. SIRT1 may therefore function as a gatekeeper of the downstream mTOR-HIF1α axis (Figure 2). Furthermore, mTOR-HIF1α-IL-9 promoter transcriptional regulation coupled with modulation of glycolytic activity is selective for SIRT1-dependent TH9 cell differentiation [51].
Transcriptional factors downstream of IL-2 are critical for TH9 cell differentiation [24], and IL-2 deficient CD4+T cells do not produce IL-9. STAT5, a downstream target of IL-2, directly binds to the Il9 locus and thus promotes TH9 cell differentiation. Mechanistic studies suggest that IL-2-STAT5 signaling inhibits B cell lymphoma 6 (Bcl6) expressions and TH17 cell generation, thereby promoting TH9 cell differentiation [24, 42].
The transcription factors NF-kB and NFAT also modulate TH9 cell differentiation. Ligation of OX40 triggers sustained activation of the non-canonical NF-kB pathway in CD4+T cells during TH9 cell differentiation [35, 36]. The non-canonical transcription factor NF-kB (RelB) directly binds to the Il9 promoter region and triggers Il9 transcription under TH9-inducing conditions. The non-canonical alternative NF-kB pathway probably also acts together with other factors to promote TH9 differentiation, suggesting that it restricts the capacity of NF-kB to interact with other transcription factors at the Il9 locus. NFAT1 (nuclear factor of activated T cells) is also required together with NF-kB for IL-9 production in CD4+T cells [52]. NFAT1 alters histone modifications and chromatin structure and restricts RelA access to the Il9 promoter region.
Transcription factors control the secretion of specific cytokines that direct T cell differentiation and differentiating T cells integrate multiple, and sometimes conflicting, signals as they differentiate into particular subsets. This is especially true for TH9 cell differentiation, during which cells integrate the Treg-inducing TGF-β signal and the TH2-inducing IL-4 signal to develop a specific TH9 cell phenotype. TGF-β signaling induces Foxp3, which is a negative regulator of TH9 cell differentiation; ectopic Foxp3 expression reduces IL-9 production in TH9 cells [35, 46]. The IL-4 signal targets multiple genes and induces expression of the transcriptional factors IRF4, GATA3, and STAT6. IRF4 is essential for TH2 and TH17, but not TH9, cell differentiation. GATA3 and STAT6 are also expressed in TH2 cells. Thus, each of these transcriptional factors likely plays integral roles in Il9 gene expression and production in CD4+T cells. Therefore, these transcription factors may not directly involved in the transcriptional regulation of the Il9 gene, but act rather as molecules downregulating negative factors during TH9 differentiation, such as Foxp3, or upregulating positive factors, such as IRF4, GATA3 and STAT6, during TH9 cell differentiation.
TH9 cell differentiation and function in various diseases
TH9 cells have protective or pathological roles in several clinical diseases, including allergic diseases, autoimmune diseases, ulcerative colitis, infection with various pathogens, and anti-tumor immunity (Figure 3 and 4).
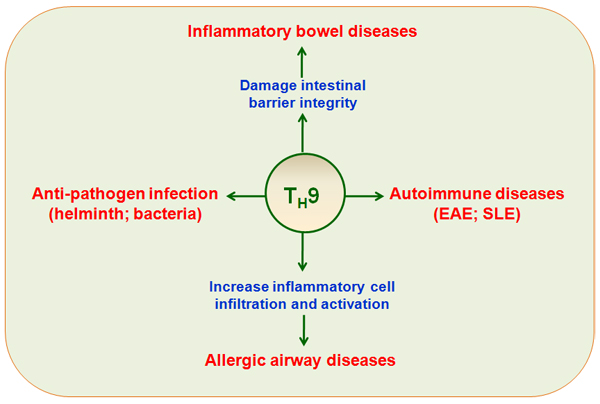
Figure 3: TH9 cells in immune-mediated diseases. TH9 cells have potent protective or pathological roles in immune-mediated diseases, including allergic airway disease, inflammatory bowel diseases, autoimmune diseases (EAE, SLE), and pathogen infections.
TH9 cells in allergic airway diseases
Allergic airway diseases are characterized by TH2-associated cytokine responses to inhaled allergens, which are orchestrated by CD4+T cells that produce eosinophilic lung inflammation [14]. In addition to the TH2-associated cytokines IL-4, IL-5, and IL-13, the TH9-associated cytokines IL-9, IL-10, and IL-21 are increased in various diseases. The TH9-associated genes IL-4RA, STAT6, TGFβRII, PU.1, OX40, IL-9, IL-9R, SMAD3, and IL-33 are related to asthma [53-55]. Airway responses and inflammatory cell levels are often evaluated in OVA-sensitized mice in asthma research [56]. Such studies indicate that TH9 cells are the primary source of IL-9 in allergic airway diseases. Additionally, increased numbers of TH9 cells in draining lymph nodes (DLNs) and airways strongly correlate with allergic airway diseases. TH9 cell-derived IL-9 can exacerbate disease conditions by increasing inflammatory cell infiltration and activation within the respiratory tract. Numbers of circulating CD4+T cells and IL-9 secretion are increased in serum from allergic lung disease patients compared to controls [57, 58]. Importantly, adoptive TH9 cell transfers increased the development of allergic airway diseases following OVA challenge, while anti-IL-9 treatment reduced disease severity [34, 58-60]. These data suggest that TH9 cells are critical for the induction of allergic airway disease. Similarly, our work showed that severe pathogenic lung inflammation and inflammatory cell recruitment, including eosinophil infiltration in bronchoalveolar lavage fluid (BALF), were increased in SIRT1-deficient mice compared to WT mice. Additionally, more of the CD4+T cells isolated from lung DLNs of SIRT1-deficient mice were IL-9+. Importantly, administration of an anti-IL-9 antibody reversed pathological lung tissue damage and infiltration of inflammatory cells, including eosinophils and IL-9+CD4+T cells in SIRT1-deficient mice. Thus, SIRT1 is required for the suppression of TH9 cell differentiation and alleviates TH9-associated allergic airway inflammation [51].
TH2 and TH9 cells may play different roles in allergic airway diseases. We propose that, during the acute inflammation phase, activated TH2 cells secrete multiple cytokines (e.g. IL-4, IL-13). Subsequently, various cytokines promote mast cell recruitment (IL-4, IL-9, and IL-13), eosinophil maturation (IL-5 and GM-CSF), basophil infiltration (IL-4), and the initiation of mucus metaplasia (IL-13). During the chronic phase of inflammation, repeated exposure to allergenic antigens and damaged epithelium promotes the secretion of various cytokine growth factors, including TGFβ, from epithelial cells, eosinophils, macrophages, and even mast cells. In the presence of TGFβ and IL-4, TH2 cells differentiate into TH9 cells, which promote the progression of allergic airway diseases [56, 61, 62].
TH9 cells in inflammatory bowel diseases
Inflammatory bowel diseases (IBD), which include Crohn’s diseases (CD) and ulcerative colitis (UC), are characterized by sustained inflammation, mucosal barrier defects, and frequent intestinal infections. TH17 and iTreg cells have been implicated in the pathological mechanisms of IBD. However, studies have also shown that TH9 cells are involved in IBD [63-65]. For example, adoptive transfer of TH9 cells into lymphopenic recipient mice (Rag1-/-; T and B cell-deficient mice) aggravated colitis, but the underlying mechanisms remain unknown. TH9 cells contribute to the pathogenesis of IBD, particularly UC, by regulating intestinal barrier integrity and immunological function [21, 66, 67]. Furthermore, IL-9 expression is elevated in patients with active UC and is highest in patients with the most severe disease. Patients with active UC also have more intestinal CD4+PU.1+T cells than control patients or patients with CD, indicating that TH9 cells are at least partly responsible for IBD pathogenesis. In acute and chronic IBD mouse models, numbers of IL-9-producing cells and IL-9R levels increased during colitis. In a novel IL-9 reporter mouse, CD4+T cells, but not other cell types such as innate lymphoid cells, are the principal source of IL-9 in colitis. Importantly, treatment with an IL-9 antibody or IL-9 knockout in mice considerably reduces colitis, as indicated by weight loss, generation of reactive oxygen species, and clinical scores, indicating that IL-9 promotes the progression of colitis. Thus, targeting IL-9 activity may improve diagnosis and treatment in UC patients.
TH9 cells in autoimmune diseases
TH9 cells have been implicated in conditions that involve central nervous system inflammation, such as systemic lupus erythematosus (SLE), experimental autoimmune encephalitis (EAE), and systemic sclerosis (SSc) [19]. Adoptive transfer of MBP (myelin basic protein)-specific TCR transgenic TH9 cells into Rag1-/- mice result in more severe EAE than transfer of TH1 or regular TH9 cells, indicating that TH9 cells play a critical role in the development of EAE [19, 68-70]. Due to complex regulatory mechanisms and variation in IL-9R singling in different immune cells during EAE, some studies have obtained conflicting results regarding the effects of IL-9 signaling in EAE [18, 43, 71]. IL-9-/- mice develop less severe EAE than their WT counterparts following either immunization with myelin proteolipid protein (PLP; 180-199) peptide in the presence of Complete Freund’s Adjuvant (CFA) or adoptive transfer of PLP (180-199) peptide-specific effector T cells from WT littermates. EAE-resistant IL-9-/- mice exhibited considerably fewer infiltrating immune cells in the CNS, as well as reduced IL-17 and IFNγ expression. IL-9 deficiency also reduced PLP peptide-specific IL-17 and IFNγ levels [72, 73]. In addition, IL-9 receptor deficiency and IL-9 neutralization attenuated EAE and correlated with decreased numbers of TH17 cells and IL-6-producing macrophages in the central nervous system, and with decreased numbers of mast cells in regional lymph nodes [74]. However, some studies found that IL-9R−/− EAE mice had more TH1 cells and TH17 cells than WT mice. Additionally, the inhibitory activity of Treg cells was reduced in IL-9R−/− mice compared to WT mice [71]. Together, these data suggest that IL-9 signaling may have a protective effect in EAE, although differential expression of IL-9 receptors in multiple cell types may result in more heterogeneous effects. However, the attenuation of EAE that results from IL-9 neutralization indicates that IL-9 signaling may also be pro-inflammatory.
To further investigate these possibilities, mRNA and serum IL-9 levels were assessed in the peripheral blood of SLE patients and healthy controls [75]. The percentage of CD4+IL-9+ T cells was elevated in SLE patients. Moreover, IL-9 expression in T cells and serum IL-9 levels in 8 untreated active SLE patients decreased 1, 2, and 3 weeks after treatment with methylprednisolone [75]. Thus, TH9 plays an important role in the pathogenesis of SLE.
TH9 cells in anti-pathogen infection activity
IL-9 production in TH9 cells also results in anti-helminth activity. Experiments in IL-9-/- mice and IL-9-fluorescent reporter mice indicate that IL-9 is crucial in the initiation of host-protective responses in the early type 2 immune response against Nippostrongylusbrasiliensis [15]. Adoptive transfer of TH9 cells, but not TH2 cells, caused rapid worm expulsion and marked basophilia and increased mast cell numbers in Rag2-/- mice [15]. TH9 cells and IL-9 are also involved in human Echinococcusgranulosus infection [76]. Compared to healthy controls, PU.1, IL-9, and GATA-3 mRNA expression were increased in untreated patients. In addition, an increase in TH9 cells with the effector memory cell phenotype was found in tuberculous pleural effusion [16]. Taken together, these data suggest that TH9 cells and IL-9 a play critical and non-redundant roles in host-protective immunity against pathogenic infection.
TH9 cell differentiation and function in anti-tumor immunity
TH9 cells regulate tumor immunity
TH9 cells are present in metastatic pleural effusion [77] and human melanoma tumor-infiltrating lymphocytes [78]. TH9 also has anti-tumor properties in mice [77, 79] (Figure 4). In addition, melanoma tumor growth is accelerated in IL-9R-/- mice, and treatment with rIL-9 inhibited this growth. Interestingly, adoptive transfer of anti-tumor TH9 cells inhibited tumor growth, and administration of an anti-IL-9 mAb reversed this effect. Moreover, treatment with exogenous IL-9 suppressed the growth of B16F10 melanoma and LLC-1, but not EL-4, tumors. Mechanistic investigation revealed that IL-9R expression is negligible in B16F10 and LLC-1 cells, and IL-9 had minimal effects on growth in these cells [80, 81]. However, IL-9R expression is increased in EL-4 cells, indicating an additional effect of IL-9 on tumor cells; evaluation of IL-9R expression may therefore be critical for IL-9 anti-tumor therapy. These results have been confirmed [77] and show that adoptive transfer of antigen-specific TH9 cells exerts anti-tumor effects by inhibiting the subcutaneous lung metastasis of B16F10 and TH9 cells. In addition, Treg, TH17, and TH2 cells also secrete low levels of IL-9. A considerable portion of TH9 cells acquire the TH1 phenotype and produce IFN-γ in vivo [82, 83]. TH9 cell plasticity therefore plays a role in various pathological processes, including cancer.
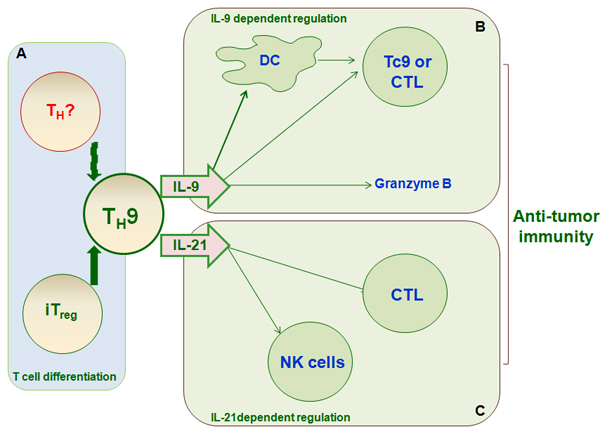
Figure 4: TH9 differentiation and function in anti-tumor immunity. TH9 cells have anti-tumor activity, particularly in melanoma. A., iTreg cells and other T cell subsets can differentiate into TH9 cells, which affect anti-tumor immunity. TH9 cells exert anti-tumor effects primarily via the secretion of the cytokines IL-9 B. and IL-21 C. IL-9 promotes tumor cell death directly by increasing granzyme B release and indirectly by increasing DC survival and recruitment, inducing host anti-tumor CD8+CTL response, or increasing Tc9 activity. IL-21 contributes to anti-tumor immunity by promoting NK cell cytolytic function and CD8+CTL response.
Our studies also indicate that TH9 cells contribute to anti-tumor immunity. Naïve T cells isolated from WT or SIRT1-deficient mice were differentiated under TH9-inducing conditions and transferred into Rag1-/- mice, which were then subcutaneously injected with B16 melanoma cells. The mice that received SIRT1-deficient CD4+T cells developed smaller tumors than WT controls. Tumor infiltrating CD4+T cells isolated from the SIRT1-deficient groups had a higher IL-9+ ratio than the WT group. Importantly, administration of an anti-IL-9 antibody reversed the changes induced by SIRT1-deficient TH9 cell transfer [51]. These data show that SIRT1 is required for the suppression of TH9 differentiation, and thus inhibits TH9-dependent anti-tumor immunity.
Regulatory mechanisms of TH9 cells in anti-tumor immunity
IL-9 produced by TH9 cells elicits anti-tumor immune responses through both direct and indirect regulatory mechanisms.
Several studies have reported direct regulatory mechanisms of TH9 cells. For example, IL-9 produced by CD4+TCells inhibits melanoma HTB-72 cell growth by upregulating p21 and TRAIL [84]. Most blood- and tissue-derived human memory TH9 cells were skin-tropic or skin-resident and co-expressed TNFα and granzyme B, suggesting that they play a pro-inflammatory role [85]. Two recent studies [86, 87] also demonstrated that glucocorticoid-induced TNF receptor-related protein (GITR) ligation directs the differentiation of iTreg cells into TH9 cells and thus mediates anti-tumor immunity, indicating that reciprocal differentiation of T cell lineages is critical for the anti-tumor effects of TH9 cells. Cca cell extract (CT26 cells) acts as an antigen and induces TH2 responses in Cca-bearing mice, thus inhibiting tumor growth primarily by converting intra-Cca Tregs cells into TH9 cells [12]. Finally, TH9 cells have stronger anti-tumor effects than TH17 or TH1 cells [22], but the reciprocal differentiation mechanisms in these T cell lineages and their effects on tumor response require further investigation.
Indirect regulatory mechanisms of TH9 cells are likely also critical for anti-tumor immunity. IL-9 produced by CD4+T cells is responsible for increased survival and function in myeloid DCs, which contribute to anti-tumor immunity [88]. TH9 cells also induce host anti-tumor CD8+CTL responses by upregulating the CCL20/CCR6-dependent recruitment of dendritic cells (DCs) to local tumor tissues [77]. These findings suggest that TH9-mediated immune regulation is crucial for anti-tumor immunity. Moreover, tumor-specific IL-9-producing CD8+Tc9 cells also exert potent anti-tumor effects [89]. Tc9 cells primed by TH9-inducing conditions secreted different cytokines and were less cytolytic in vitro, but exhibited much stronger anti-tumor effects in OT-I/B16-OVA and Pmel-1/B16 melanoma mouse models. Adoptive transfer of Tc9 cells results in differentiation into IFNγ- and granzyme-B (GrzB)-producing cytolytic Tc1-like effector cells. This suggests that the anti-tumor activities of TH9 cells are partially due to their effects on Tc9 cells.
In addition to IL-9, IL-21 secreted by TH9 cells also exerts critical anti-tumor effects. IL-1β promotes the secretion of cytokines IL-9, IL-10, and IL-21 from TH9 cells through STAT1-IRF1-dependent mechanisms [22, 90, 91]. Furthermore, this IL-1β-induced TH9 differentiation increased IL-21, but not IL-9, secretion, and increased IL-21-dependent anti-tumor effects [92-94]. IL-21 stimulates IFNγ production, enhances the cytolytic activity of NK cells, and increases CD8+CTL activity, all of which promote anti-tumor immunity [28, 94].
Concluding remarks
Following antigen stimulation, naïve CD4+T cells differentiate into one of several functional effector cell classes. In addition to the classical TH1 and TH2 lineages, TH17 cells have been extensively characterized. Recently, new subsets of IL-9-producing CD4+T cells have been induced in vitro by IL-4 and TGFβ and identified in vivo. IL-9 is a pleiotropic cytokine produced by TH2 cells, mast cells, and eosinophils. Due to the critical role of IL-9-producing CD4+T cells in immune-mediated diseases, TH9 cells have been extensively investigated in mouse and human studies during the past 20 years. These studies have demonstrated that TH9 cells contribute to both immune responses and immunopathological diseases. Moreover, they suggest that promoting or suppressing TH9 cell differentiation, transcriptional control, and function may provide novel treatments for TH9-associated inflammation, autoimmune diseases, and tumors. Specifically, administration of IL-9 antibodies effectively eliminated TH9 cells and ameliorated or aggravatedTH9-associated diseases.
Abbreviations
BCL6: B cell lymphoma 6;
CD: Crohn’s diseases;
DCs: dendritic cells;
EAE: experimental autoimmune encephalitis;
IL-9: interleukin 9;
IBD: Inflammatory bowel diseases;
iTreg cells: inducible regulatory T cells;
HAT: histone acetyltransferase;
NFAT: nuclear factor of activated T cells;
NK cells: natural killing cells;
NF-kB: nuclear factor-kappa B;
NICD: notch intracellular domain;
RBP-Jk: recombination signal binding protein for immunoglobulin kappa J region;
STAT6: signal transducer and activator of transcription 6;
SLE: systemic lupus erythematosus;
SSc: systemic sclerosis;
TNFα: tumor necrosis factorα;
TSLP: thymic stromal lymphopoietin;
TGFβ1: transforming growth factor β1;
TH9: IL-9 producing CD4+T cells;
GITR: TNF receptor-related protein;
GrzB: granzyme-B;
UC: ulcerative colitis
Conflicts of interest
The authors declare no competing financial interests.
Grant support
This research was supported by grants from the National Natural Science Foundation for General Programs of China (31671524, 31171407 and 81273201, G.L.), the Key Basic Research Project of the Science and Technology Commission of Shanghai Municipality (12JC1400900, G.L.), and the Innovation Program of the Shanghai Municipal Education Commission (14ZZ009, G.L.).
References
1. Kaplan MH, Hufford MM and Olson MR. The development and in vivo function of T helper 9 cells. Nat Rev Immunol. 2015; 15: 295-307.
2. Powell JD and Delgoffe GM. The mammalian target of rapamycin: linking T cell differentiation, function, and metabolism. Immunity. 2010; 33: 301-311.
3. Yosef N, Shalek AK, Gaublomme JT, Jin H, Lee Y, Awasthi A, Wu C, Karwacz K, Xiao S, Jorgolli M, Gennert D, Satija R, Shakya A, et al. Dynamic regulatory network controlling TH17 cell differentiation. Nature. 2013; 496: 461-468.
4. Ivanov, II, Zhou L and Littman DR. Transcriptional regulation of Th17 cell differentiation. Semin Immunol. 2007; 19: 409-417.
5. Liu G, Burns S, Huang G, Boyd K, Proia RL, Flavell RA and Chi H. The receptor S1P1 overrides regulatory T cell-mediated immune suppression through Akt-mTOR. Nat Immunol. 2007; 10: 769-777.
6. Liu G, Bi Y, Xue L, Zhang Y, Yang H, Chen X, Lu Y, Zhang Z, Liu H, Wang X, Wang R, Chu Y and Yang R. Dendritic cell SIRT1-HIF1alpha axis programs the differentiation of CD4+ T cells through IL-12 and TGF-beta1. Proc Natl Acad Sci U S A. 2015; 112: E957-965.
7. Bi Y, Liu G and Yang R. Reciprocal modulation between TH17 and other helper T cell lineages. J Cell Physiol. 2011; 226: 8-13.
8. Bi Y, Liu G and Yang R. Th17 cell induction and immune regulatory effects. J Cell Physiol. 2007; 211: 273-278.
9. Schmitt E, Germann T, Goedert S, Hoehn P, Huels C, Koelsch S, Kuhn R, Muller W, Palm N and Rude E. IL-9 production of naive CD4+ T cells depends on IL-2, is synergistically enhanced by a combination of TGF-beta and IL-4, and is inhibited by IFN-gamma. J Immunol. 1994; 153: 3989-3996.
10. Dardalhon V, Awasthi A, Kwon H, Galileos G, Gao W, Sobel RA, Mitsdoerffer M, Strom TB, Elyaman W, Ho IC, Khoury S, Oukka M and Kuchroo VK. IL-4 inhibits TGF-beta-induced Foxp3+ T cells and, together with TGF-beta, generates IL-9+ IL-10+ Foxp3(-) effector T cells. Nat Immunol. 2008; 9: 1347-1355.
11. Veldhoen M, Uyttenhove C, van Snick J, Helmby H, Westendorf A, Buer J, Martin B, Wilhelm C and Stockinger B. Transforming growth factor-beta ‘reprograms’ the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat Immunol. 2008; 9: 1341-1346.
12. Liu JQ, Li XY, Yu HQ, Yang G, Liu ZQ, Geng XR, Wang S, Mo LH, Zeng L, Zhao M, Fu YT, Sun HZ, Liu ZG, et al. Tumor-specific Th2 responses inhibit growth of CT26 colon-cancer cells in mice via converting intratumor regulatory T cells to Th9 cells. Sci Rep. 2015; 5:10665.
13. Maier E, Werner D, Duschl A, Bohle B and Horejs-Hoeck J. Human Th2 but not Th9 cells release IL-31 in a STAT6/NF-kappaB-dependent way. J Immunol. 2014; 193: 645-654.
14. Froidure A, Shen C, Gras D, Van Snick J, Chanez P and Pilette C. Myeloid dendritic cells are primed in allergic asthma for thymic stromal lymphopoietin-mediated induction of Th2 and Th9 responses. Allergy. 2014; 69: 1068-1076.
15. Licona-Limon P, Henao-Mejia J, Temann AU, Gagliani N, Licona-Limon I, Ishigame H, Hao L, Herbert DR and Flavell RA. Th9 cells drive host immunity against gastrointestinal worm infection. Immunity. 2013; 39:744-57.
16. Ye ZJ, Yuan ML, Zhou Q, Du RH, Yang WB, Xiong XZ, Zhang JC, Wu C, Qin SM and Shi HZ. Differentiation and recruitment of Th9 cells stimulated by pleural mesothelial cells in human Mycobacterium tuberculosis infection. PLoS One. 2012; 7:e31710.
17. Kara EE, Comerford I, Bastow CR, Fenix KA, Litchfield W, Handel TM and McColl SR. Distinct chemokine receptor axes regulate Th9 cell trafficking to allergic and autoimmune inflammatory sites. J Immunol. 2013; 191:1110-1117.
18. Murugaiyan G, Beynon V, Pires Da Cunha A, Joller N and Weiner HL. IFN-gamma limits Th9-mediated autoimmune inflammation through dendritic cell modulation of IL-27. J Immunol. 2012; 189:5277-5283.
19. Jager A, Dardalhon V, Sobel RA, Bettelli E and Kuchroo VK. Th1, Th17, and Th9 effector cells induce experimental autoimmune encephalomyelitis with different pathological phenotypes. J Immunol. 2009; 183:7169-7177.
20. Leake I. IBD. TH9 cells might have a role in the pathogenesis of ulcerative colitis. Nat Rev Gastroenterol Hepatol. 2014; 11:455-523.
21. Gerlach K, Hwang Y, Nikolaev A, Atreya R, Dornhoff H, Steiner S, Lehr HA, Wirtz S, Vieth M, Waisman A, Rosenbauer F, McKenzie AN, Weigmann Bet al. TH9 cells that express the transcription factor PU.1 drive T cell-mediated colitis via IL-9 receptor signaling in intestinal epithelial cells. Nat Immunol. 2014; 15:676-686.
22. Vegran F, Apetoh L and Ghiringhelli F. Th9 cells: a novel CD4 T-cell subset in the immune war against cancer. Cancer Res. 2015; 75:475-479.
23. Anuradha R, George PJ, Hanna LE, Chandrasekaran V, Kumaran P, Nutman TB and Babu S. IL-4-, TGF-beta-, and IL-1-dependent expansion of parasite antigen-specific Th9 cells is associated with clinical pathology in human lymphatic filariasis. J Immunol. 2013; 191:2466-2473.
24. Liao W, Spolski R, Li P, Du N, West EE, Ren M, Mitra S and Leonard WJ. Opposing actions of IL-2 and IL-21 on Th9 differentiation correlate with their differential regulation of BCL6 expression. Proc Natl Acad Sci U S A. 2014; 111:3508-3513.
25. Gomez-Lopez N, Olson DM and Robertson SA. Interleukin-6 controls uterine Th9 cells and CD8(+) T regulatory cells to accelerate parturition in mice. Immunol Cell Biol. 2016; 94:79-89.
26. Anuradha R, Munisankar S, Bhootra Y, Jagannathan J, Dolla C, Kumaran P, Nutman TB and Babu S. IL-10- and TGFbeta-mediated Th9 Responses in a Human Helminth Infection. PLoS Negl Trop Dis. 2016; 10:e0004317.
27. Wong MT, Ye JJ, Alonso MN, Landrigan A, Cheung RK, Engleman E and Utz PJ. Regulation of human Th9 differentiation by type I interferons and IL-21. Immunol Cell Biol. 2010; 88:624-631.
28. Vegran F, Berger H, Boidot R, Mignot G, Bruchard M, Dosset M, Chalmin F, Rebe C, Derangere V, Ryffel B, Kato M, Prevost-Blondel A, Ghiringhelli F, et al. The transcription factor IRF1 dictates the IL-21-dependent anticancer functions of TH9 cells. Nat Immunol. 2014; 15:758-766.
29. Guo W, Yu D, Wang X, Luo C, Chen Y, Lei W, Wang C, Ge Y, Xue W, Tian Q, Gao X and Yao W. Anti-inflammatory effects of Interleukin-23 receptor cytokine-binding homology region rebalance T cell distribution in rodent collagen-induced arthritis. Oncotarget. 2016; 7:31800-13. doi: 10.18632/oncotarget.9309.
30. Neill DR and McKenzie AN. TH9 cell generation. TH9: the latest addition to the expanding repertoire of IL-25 targets. Immunol Cell Biol. 2010; 88:502-504.
31. Blom L, Poulsen BC, Jensen BM, Hansen A and Poulsen LK. IL-33 induces IL-9 production in human CD4+ T cells and basophils. PLoS One. 2011; 6:e21695.
32. Yao W, Zhang Y, Jabeen R, Nguyen ET, Wilkes DS, Tepper RS, Kaplan MH and Zhou B. Interleukin-9 is required for allergic airway inflammation mediated by the cytokine TSLP. Immunity. 2013; 38:360-372.
33. Nakatsukasa H, Zhang D, Maruyama T, Chen H, Cui K, Ishikawa M, Deng L, Zanvit P, Tu E, Jin W, Abbatiello B, Goldberg N, Chen Q, et al. The DNA-binding inhibitor Id3 regulates IL-9 production in CD4(+) T cells. Nat Immunol. 2015; 16:1077-1084.
34. Yang XO, Zhang H, Kim BS, Niu X, Peng J, Chen Y, Kerketta R, Lee YH, Chang SH, Corry DB, Wang D, Watowich SS and Dong C. The signaling suppressor CIS controls proallergic T cell development and allergic airway inflammation. Nat Immunol. 2013; 14:732-740.
35. Goswami R and Kaplan MH. Yoking OX40 to regulation of IL-9. Nat Immunol. 2012; 13:942-943.
36. Xiao X, Balasubramanian S, Liu W, Chu X, Wang H, Taparowsky EJ, Fu YX, Choi Y, Walsh MC and Li XC. OX40 signaling favors the induction of T(H)9 cells and airway inflammation. Nat Immunol. 2012; 13:981-990.
37. Demoulin JB, Uyttenhove C, Van Roost E, DeLestre B, Donckers D, Van Snick J and Renauld JC. A single tyrosine of the interleukin-9 (IL-9) receptor is required for STAT activation, antiapoptotic activity, and growth regulation by IL-9. Mol Cell Biol. 1996; 16:4710-4716.
38. Goswami R and Kaplan MH. Gcn5 is required for PU.1-dependent IL-9 induction in Th9 cells. J Immunol. 2012; 189:3026-3033.
39. Chang HC, Sehra S, Goswami R, Yao W, Yu Q, Stritesky GL, Jabeen R, McKinley C, Ahyi AN, Han L, Nguyen ET, Robertson MJ, Perumal NB, et al. The transcription factor PU.1 is required for the development of IL-9-producing T cells and allergic inflammation. Nat Immunol. 2010; 11:527-534.
40. Tamiya T, Ichiyama K, Kotani H, Fukaya T, Sekiya T, Shichita T, Honma K, Yui K, Matsuyama T, Nakao T, Fukuyama S, Inoue H, Nomura M, et al. Smad2/3 and IRF4 play a cooperative role in IL-9-producing T cell induction. J Immunol. 2013; 191:2360-2371.
41. Stassen M, Muller C, Arnold M, Hultner L, Klein-Hessling S, Neudorfl C, Reineke T, Serfling E and Schmitt E. IL-9 and IL-13 production by activated mast cells is strongly enhanced in the presence of lipopolysaccharide: NF-kappa B is decisively involved in the expression of IL-9. J Immunol. 2001; 166:4391-4398.
42. Bassil R, Orent W, Olah M, Kurdi AT, Frangieh M, Buttrick T, Khoury SJ and Elyaman W. BCL6 controls Th9 cell development by repressing Il9 transcription. J Immunol. 2014; 193:198-207.
43. Elyaman W, Bassil R, Bradshaw EM, Orent W, Lahoud Y, Zhu B, Radtke F, Yagita H and Khoury SJ. Notch receptors and Smad3 signaling cooperate in the induction of interleukin-9-producing T cells. Immunity. 2012; 36:623-634.
44. Ramming A, Druzd D, Leipe J, Schulze-Koops H and Skapenko A. Maturation-related histone modifications in the PU.1 promoter regulate Th9-cell development. Blood. 2012; 119:4665-4674.
45. Jabeen R and Kaplan MH. The symphony of the ninth: the development and function of Th9 cells. Curr Opin Immunol. 2012; 24:303-307.
46. Goswami R, Jabeen R, Yagi R, Pham D, Zhu J, Goenka S and Kaplan MH. STAT6-dependent regulation of Th9 development. J Immunol. 2012; 188:968-975.
47. Li S, Guo X, Lu LF, Lu XB, Wu N and Zhang YA. Regulation pattern of fish irf4 (the gene encoding IFN regulatory factor 4) by STAT6, c-Rel and IRF4. Dev Comp Immunol. 2015; 51:65-73.
48. Amsen D, Antov A, Jankovic D, Sher A, Radtke F, Souabni A, Busslinger M, McCright B, Gridley T and Flavell RA. Direct regulation of Gata3 expression determines the T helper differentiation potential of Notch. Immunity. 2007; 27:89-99.
49. Fang TC, Yashiro-Ohtani Y, Del Bianco C, Knoblock DM, Blacklow SC and Pear WS. Notch directly regulates Gata3 expression during T helper 2 cell differentiation. Immunity. 2007; 27:100-110.
50. Beckebaum S, Kabar I and Cicinnati VR. Hepatitis B and C in liver transplantation: new strategies to combat the enemies. Rev Med Virol. 2013; 23:172-193.
51. Wang Y, Bi Y, Chen X, Li C, Li Y, Zhang Z, Wang J, Lu Y, Yu Q, Su H, Yang H, Liu G. Histone Deacetylase SIRT1 negatively regulates the differentiation of interleukin 9- producing CD4+T cells. Immunity. 2016; 44:1337-1349.
52. Jash A, Sahoo A, Kim GC, Chae CS, Hwang JS, Kim JE and Im SH. Nuclear factor of activated T cells 1 (NFAT1)-induced permissive chromatin modification facilitates nuclear factor-kappaB (NF-kappaB)-mediated interleukin-9 (IL-9) transactivation. J Biol Chem. 2012; 287:15445-15457.
53. Simpson LJ, Patel S, Bhakta NR, Choy DF, Brightbill HD, Ren X, Wang Y, Pua HH, Baumjohann D, Montoya MM, Panduro M, Remedios KA, Huang X, et al. A microRNA upregulated in asthma airway T cells promotes TH2 cytokine production. Nat Immunol. 2014; 15:1162-1170.
54. Seumois G, Chavez L, Gerasimova A, Lienhard M, Omran N, Kalinke L, Vedanayagam M, Ganesan AP, Chawla A, Djukanovic R, Ansel KM, Peters B, Rao A, et al. Epigenomic analysis of primary human T cells reveals enhancers associated with TH2 memory cell differentiation and asthma susceptibility. Nat Immunol. 2014; 15:777-788.
55. Parulekar AD, Atik MA and Hanania NA. Periostin, a novel biomarker of TH2-driven asthma. Curr Opin Pulm Med. 2014; 20:60-65.
56. Holt PG and Sly PD. Th2 cytokines in the asthma late-phase response. Lancet. 2007; 370:1396-1398.
57. Kudo M, Ishigatsubo Y and Aoki I. Pathology of asthma. Front Microbiol. 2013; 4:263-272.
58. Soroosh P and Doherty TA. Th9 and allergic disease. Immunology. 2009; 127:450-458.
59. Staudt V, Bothur E, Klein M, Lingnau K, Reuter S, Grebe N, Gerlitzki B, Hoffmann M, Ulges A, Taube C, Dehzad N, Becker M, Stassen M, et al. Interferon-regulatory factor 4 is essential for the developmental program of T helper 9 cells. Immunity. 2010; 33:192-202.
60. Boyce JA and Peebles RS. Regulation of Th9-type pulmonary immune responses: a new role for COX-2. Am J Respir Crit Care Med. 2013; 187:785-786.
61. Maziak W. The Th1-Th2 paradigm and asthma: how far should we go? J Asthma. 2003; 40:201-215.
62. Komai M, Tanaka H, Masuda T, Nagao K, Ishizaki M, Sawada M and Nagai H. Role of Th2 responses in the development of allergen-induced airway remodelling in a murine model of allergic asthma. Br J Pharmacol. 2003; 138:912-920.
63. Ciucci T, Ibanez L, Boucoiran A, Birgy-Barelli E, Pene J, Abou-Ezzi G, Arab N, Rouleau M, Hebuterne X, Yssel H, Blin-Wakkach C and Wakkach A. Bone marrow Th17 TNFalpha cells induce osteoclast differentiation, and link bone destruction to IBD. Gut. 2015; 64:1072-1081.
64. Wu W, He C, Liu C, Cao AT, Xue X, Evans-Marin HL, Sun M, Fang L, Yao S, Pinchuk IV, Powell DW, Liu Z and Cong Y. miR-10a inhibits dendritic cell activation and Th1/Th17 cell immune responses in IBD. Gut. 2015; 64:1755-1764.
65. Galvez J. Role of Th17 Cells in the Pathogenesis of Human IBD. ISRN Inflamm 2014:928461.
66. Gerlach K, McKenzie AN, Neurath MF and Weigmann B. IL-9 regulates intestinal barrier function in experimental T cell-mediated colitis. Tissue Barriers. 2015; 3:e983777.
67. Nalleweg N, Chiriac MT, Podstawa E, Lehmann C, Rau TT, Atreya R, Krauss E, Hundorfean G, Fichtner-Feigl S, Hartmann A, Becker C and Mudter J. IL-9 and its receptor are predominantly involved in the pathogenesis of UC. Gut. 2015; 64:743-755.
68. Pan HF, Leng RX, Li XP, Zheng SG and Ye DQ. Targeting T-helper 9 cells and interleukin-9 in autoimmune diseases. Cytokine Growth Factor Rev. 2013; 24:515-522.
69. Sandy AR, Stoolman J, Malott K, Pongtornpipat P, Segal BM and Maillard I. Notch signaling regulates T cell accumulation and function in the central nervous system during experimental autoimmune encephalomyelitis. J Immunol. 2013; 191:1606-1613.
70. Rostami A and Ciric B. Role of Th17 cells in the pathogenesis of CNS inflammatory demyelination. J Neurol Sci. 2013; 333:76-87.
71. Elyaman W, Bradshaw EM, Uyttenhove C, Dardalhon V, Awasthi A, Imitola J, Bettelli E, Oukka M, van Snick J, Renauld JC, Kuchroo VK and Khoury SJ. IL-9 induces differentiation of TH17 cells and enhances function of FoxP3+ natural regulatory T cells. Proc Natl Acad Sci U S A. 2009; 106:12885-91280.
72. Li H, Nourbakhsh B, Cullimore M, Zhang GX and Rostami A. IL-9 is important for T-cell activation and differentiation in autoimmune inflammation of the central nervous system. Eur J Immunol. 2011; 41:2197-2206.
73. Li H, Nourbakhsh B, Ciric B, Zhang GX and Rostami A. Neutralization of IL-9 ameliorates experimental autoimmune encephalomyelitis by decreasing the effector T cell population. J Immunol. 2010; 185:4095-4100.
74. Nowak EC, Weaver CT, Turner H, Begum-Haque S, Becher B, Schreiner B, Coyle AJ, Kasper LH and Noelle RJ. IL-9 as a mediator of Th17-driven inflammatory disease. J Exp Med. 2009; 206:1653-1660.
75. Ouyang H, Shi Y, Liu Z, Feng S, Li L, Su N, Lu Y and Kong S. Increased interleukin9 and CD4+IL-9+ T cells in patients with systemic lupus erythematosus. Mol Med Rep. 2013; 7:1031-1037.
76. Pang N, Zhang F, Ma X, Zhang Z, Zhao H, Xin Y, Wang S, Zhu Y, Wen H and Ding J. Th9/IL-9 profile in human echinococcosis: their involvement in immune response during infection by Echinococcus granulosus. Mediators Inflamm. 2014:781649.
77. Lu Y, Hong S, Li H, Park J, Hong B, Wang L, Zheng Y, Liu Z, Xu J, He J, Yang J, Qian J and Yi Q. Th9 cells promote antitumor immune responses in vivo. J Clin Invest. 2012; 122:4160-4171.
78. Ye ZJ, Zhou Q, Yin W, Yuan ML, Yang WB, Xiong XZ, Zhang JC and Shi HZ. Differentiation and immune regulation of IL-9-producing CD4+ T cells in malignant pleural effusion. Am J Respir Crit Care Med. 2012; 186:1168-1179.
79. Purwar R, Schlapbach C, Xiao S, Kang HS, Elyaman W, Jiang X, Jetten AM, Khoury SJ, Fuhlbrigge RC, Kuchroo VK, Clark RA and Kupper TS. Robust tumor immunity to melanoma mediated by interleukin-9-producing T cells. Nat Med. 2012; 18:1248-1253.
80. Lv X and Wang X. The role of interleukin-9 in lymphoma. Leuk Lymphoma. 2013; 54:1367-1372.
81. Eyles J, Puaux AL, Wang X, Toh B, Prakash C, Hong M, Tan TG, Zheng L, Ong LC, Jin Y, Kato M, Prevost-Blondel A, Chow P, et al. Tumor cells disseminate early, but immunosurveillance limits metastatic outgrowth, in a mouse model of melanoma. J Clin Invest. 2010; 120:2030-2039.
82. Tan C, Aziz MK, Lovaas JD, Vistica BP, Shi G, Wawrousek EF and Gery I. Antigen-specific Th9 cells exhibit uniqueness in their kinetics of cytokine production and short retention at the inflammatory site. J Immunol. 2010; 185:6795-6801.
83. Umezu-Goto M, Kajiyama Y, Kobayashi N, Kaminuma O, Suko M and Mori A. IL-9 production by peripheral blood mononuclear cells of atopic asthmatics. Int Arch Allergy Immunol. 2007; 143 Suppl 1:76-79.
84. Fang Y, Chen X, Bai Q, Qin C, Mohamud AO, Zhu Z, Ball TW, Ruth CM, Newcomer DR, Herrick EJ and Nicholl MB. IL-9 inhibits HTB-72 melanoma cell growth through upregulation of p21 and TRAIL. J Surg Oncol. 2015; 111:969-974.
85. Schlapbach C, Gehad A, Yang C, Watanabe R, Guenova E, Teague JE, Campbell L, Yawalkar N, Kupper TS and Clark RA. Human TH9 cells are skin-tropic and have autocrine and paracrine proinflammatory capacity. Sci Transl Med. 2014; 6:219ra8.
86. Xiao X, Shi X, Fan Y, Zhang X, Wu M, Lan P, Minze L, Fu YX, Ghobrial RM, Liu W and Li XC. GITR subverts Foxp3(+) Tregs to boost Th9 immunity through regulation of histone acetylation. Nat Commun. 2015; 6:8266-8275.
87. Kim IK, Kim BS, Koh CH, Seok JW, Park JS, Shin KS, Bae EA, Lee GE, Jeon H, Cho J, Jung Y, Han D, Kwon BS, et al. Glucocorticoid-induced tumor necrosis factor receptor-related protein co-stimulation facilitates tumor regression by inducing IL-9-producing helper T cells. Nat Med. 2015; 21:1010-1017.
88. Park J, Li H, Zhang M, Lu Y, Hong B, Zheng Y, He J, Yang J, Qian J and Yi Q. Murine Th9 cells promote the survival of myeloid dendritic cells in cancer immunotherapy. Cancer Immunol Immunother. 2014; 63:835-845.
89. Lu Y, Hong B, Li H, Zheng Y, Zhang M, Wang S, Qian J and Yi Q. Tumor-specific IL-9-producing CD8+ Tc9 cells are superior effector than type-I cytotoxic Tc1 cells for adoptive immunotherapy of cancers. Proc Natl Acad Sci U S A. 2014; 111:2265-2270.
90. Jia L and Wu C. Differentiation, regulation and function of Th9 cells. Adv Exp Med Biol. 2014; 841:181-207.
91. Quezada SA and Peggs KS. An antitumor boost to TH9 cells. Nat Immunol. 2014; 15:703-705.
92. Petrella TM, Tozer R, Belanger K, Savage KJ, Wong R, Smylie M, Kamel-Reid S, Tron V, Chen BE, Hunder NN, Hagerman L, Walsh W and Eisenhauer EA. Interleukin-21 has activity in patients with metastatic melanoma: a phase II study. J Clin Oncol. 2012; 30:3396-3401.
93. Zeng R, Spolski R, Finkelstein SE, Oh S, Kovanen PE, Hinrichs CS, Pise-Masison CA, Radonovich MF, Brady JN, Restifo NP, Berzofsky JA and Leonard WJ. Synergy of IL-21 and IL-15 in regulating CD8+ T cell expansion and function. J Exp Med. 2005; 201:139-148.
94. Strengell M, Matikainen S, Siren J, Lehtonen A, Foster D, Julkunen I and Sareneva T. IL-21 in synergy with IL-15 or IL-18 enhances IFN-gamma production in human NK and T cells. J Immunol. 2003; 170:5464-5469.