
INTRODUCTION
In spite of the recent advancements in the multidisciplinary treatment for oral squamous cell carcinoma (OSCC), OSCC remains one of the leading causes of cancer-related mortality worldwide [1]. Extensive epidemiologic evidence have demonstrated increased risk for the development of OSCC associated with areca nut chewing in Southeast Asia [2, 3]. Mounting short-term assay studies have demonstrated that arecoline, a major areca nut alkaloid, contributes pathogenesis of OSCC [4, 5]. Nevertheless, the underlying mechanisms by which the long-term arecoline participates in tumorigenesis of OSCC are not well understood.
Recent studies have revealed that cancer stem cells (CSCs) or termed tumor initiating cells (TICs) with tumors could contribute to tumor maintenance, metastasis, radio-resistance and chemo-resistance in a variety of cancers, including OSCC [6–11]. Moreover, the first connection of between CSCs and epithelial-mesenchymal transdifferentiation (EMT), a dynamic process in which cells lose epithelial features and gain mesenchymal properties, is demonstrated in breast cancer stem cells model [12]. Mani et al. found that mammary epithelial cells treated with EMT inducer TGFβ increased cancer stemness marker [12]. Our previous report has shown that S100A4, a mediator of EMT, plays a crucial role the regulation of cancer stemness and tumorigenic properties both in vitro and in vivo [13]. Overexpression of cancer stemness marker CD133 also increased EMT transformation in OSCC [8]. Therefore, understanding the relationships between areca nut and CSCs/EMT is important to improve further OSCC therapeutics.
In this study, we developed a chronic arecoline-treated oral epithelial cells model for phenotypic and molecular characterization of the arecoline-induced cancer stemness and EMT. The aim of this study was to explore whether long-term chronic arecoline treatment positively correlated with cancer stemness and EMT in OE cells. To the best of our knowledge, we found that long-term arecoline treatment enhanced the in vitro and in vivo tumorigenicity of OE cells, which could be blocked by miR-145 delivery. This study might open a new avenue for cancer stemness generation by areca nut and to be able to develop innovative treatments for areca nut-associated OSCC patients.
RESULTS
Elevation of cancer stemness marker ADLH1 activity and CD44 positivity in long-term arecoline-exposed oral epithelial cells
The mechanism by which chronic areca nut treatment progresses to OSCC is poorly described. We hypothesize that one of the mechanisms contributing to the oral carcinogenesis involves cancer stemness enhancement. Two oral epithelial cell lines, SG and FaDu cells, were treated with arecoline up to three months for cancer stemness evaluation compared with their parental cells. Mounting reports suggested that aldehyde dehydrogenase (ALDH) activity [14] and CD44 [7] expression could be the common markers to identify the oral CSCs. Our data reported that long-term arecoline sustained treatment dose-dependently elevated ALDH1 activity of OE cells (Figure 1A). CD44 expression levels were also higher in arecoline-exposed OE cells, but lower in parental OE cells (Figure 1B).
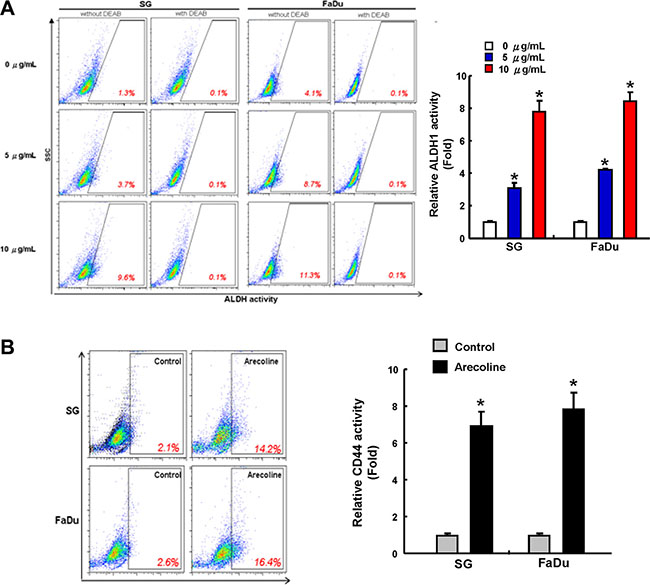
Figure 1: Increase of ALDH1 and CD44 activity in oral epithelial cells with long-term arecoline exposure. The expression of ALDH1 activity (A) and CD44 positivity (B) of control and long-term arecoline exposed oral epithelial cells (SG and FaDu) was determined by flow cytometry analysis. Data shown here are the mean ± SD of three independent experiments. *p < 0.05 vs. Control.
Chronic arecoline treatment increases sphere-forming capacity and the expression of stemness markers
Oralsphere formation over serial passages of culture is gold standard methodology for evaluating the self-renewal property in CSCs [15]. Accordingly, the spheres body size (Figure 2A), efficiency of secondary sphere/tertiary sphere formation (Figure 2B) efficiency was increased after chronic arecoline exposure. Real-time RT-PCR (Figure 2C) and western blotting analysis (Figure 2D) also showed up-regulation of stemness markers expression (Oct4, Nanog, and Sox2) in arecoline-exposed OE cells compared with their parental cells.
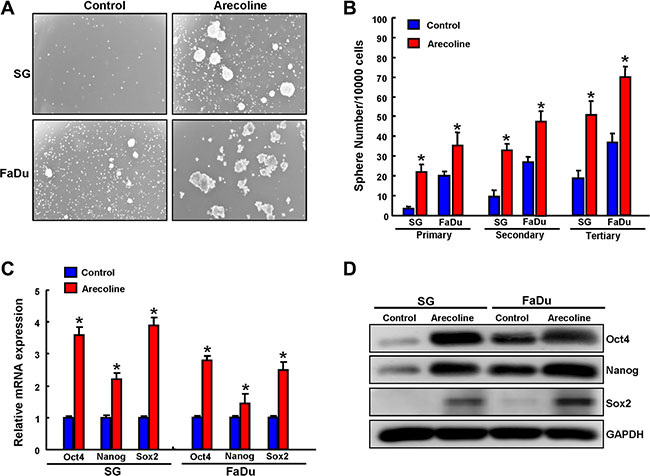
Figure 2: Enhanced self-renewal capacity and pluripotency genes in long-term arecoline-exposed oral epithelial cells. (A) Representative image of control and long-term arecoline exposed oral epithelial cells (SG and FaDu) culturing in a serum-free medium were subjected to secondary oralspheres formation assays. (B) The secondary oralspheres generated from control and long-term arecoline exposed OE cells during three passages were quantified. (C) qRT-PCR analysis and (D) Western blot analysis of Nanog, Oct4 and Sox2 expression in the control and long-term arecoline exposed OE cell subclones. Data shown here are the mean ± SD of three independent experiments. *p < 0.05 vs. Control.
Chronic arecoline-exposed oral epithelial cells showed chemoresistence to cisplatin and 5-FU
The observation of arecoline-enhanced the CSCs population and its properties suggested their involvement in modulating the chemoresistance, an important hallmark of CSCs [16]. To explore the expression of arecoline-exposed OE cells and chemoresistance, control and arecoline-exposed OE cells treated with chemotherapeutic agents and subjected to MTT analysis. MTT analysis revealed that OE cells with long-term arecoline exposure significantly increased chemoresistence to cisplatin (Figure 3A) and 5-FU (Figure 3B) compared to the parental OE cells.
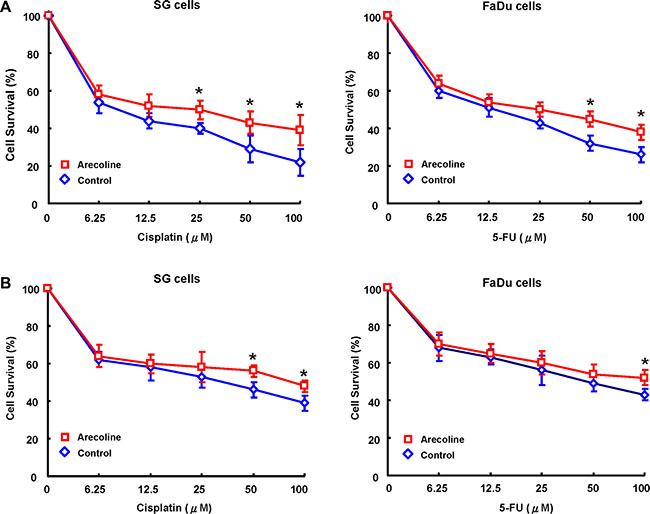
Figure 3: Long-term arecoline exposure increased chemoresistance in oral epithelial cells. Control and long-term arecoline exposed oral epithelial cells were subjected to treatment with different concentrations of cisplatin (A) and 5-FU (B). Cell viability was determined by MTT assay (*p < 0.05).
Increased tumorigenicity and EMT properties in oral epithelial cells with long-term arecoline exposure
To assess the impact of chronic arecoline exposure on tumorigenicity in vitro and in vivo, we performed migration/invasion assay using transwell system and colony-forming assay. Migration (Figure 4A), invasion (Figure 4B) and colony formation (Figure 4C) were also enhanced in long-term arecoline-exposed OE cells when compared with the corresponding untreated OE cells. Comparing to control OE cells, xenotransplantation of long-term arecoline-treated OE cells induced severe tumor formation in immunocompromised mice at 25 day post-transplantation (Figure 4D). Epithelial-mesenchymal transition (EMT), a de-differentiation program that converts adherent epithelial cells into individual migratory cells, is thought to be a cellular process commonly associated with CSCs [12]. Real-time RT-PCR analysis also demonstrated increased transcripts of mesenchymal markers and reduced epithelial marker in chronic arecoline-exposed OE (AOE) cells (Figure 4E). Consistently, AOE cells also increased protein levels of mesenchymal markers and reduced epithelial marker by western blotting (Figure 4F).
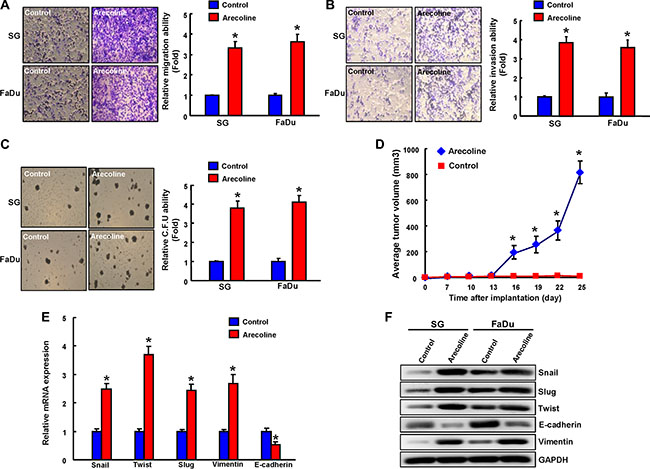
Figure 4: Enhanced in vitro and in vivo oncogenicity in long-term arecoline-stimulated oral epithelial cells. Control and long-term arecoline exposed oral epithelial cells were subjected to migration assay (A), matrix invasion assay (B), and a soft agar colony formation assay (C). (D) Xenograft tumor volume in recipients of 1 × 104 control (yellow arrow) and long-term arecoline-treated OE cells (red arrow) were monitored over 25 day experimental period. Expression of EMT-related markers in control and long-term arecoline-treated OE cells was determined by (E) qRT-PCR analysis and (F) Western blot analysis. The amount of GAPDH protein of different crude cell extracts was referred as loading control for further quantification. Data shown here are the mean ± SD of three independent experiments. *p < 0.05 vs. Control.
miR-145 ablated arecoline-induced cancer stemness and in vivo tumorigenesis
MicroRNAs (miRNAs), a class of small noncoding RNAs regulating the gene expression by binding to the 3′ untranslated region (UTR) of target mRNAs, have been involved in cancer stemness and EMT during carcinogenesis [17]. miRNAs microarray analyses identified miR-145, the known tumor suppressive microRNA, is significant down-regulated in AOE cells (Figure 5A). Consistent with the miRNA microarray results, long-term arecoline exposure dose-dependently led to the marked down-regulation of miR-145 expression by miRNA real-time RT-PCR analysis (Figure 5B). To further investigate the effect of miR-145 on biological properties of AOE cells, we gained-of function miR-145 in AOE cells through lentiviral-mediated transduction (Figure 5C). miRNA real-time RT-PCR analysis confirmed the miR-145 over-expression effect in AOE cells (Figure 5C). The sphere-forming ability among the primary, secondary and tertiary spheres was consistently impaired in AOE cells with miR-145 overexpression (Figure 5D). The formation of soft agar colonies (Figure 5E) and invasiveness ability (Figure 5F) was suppressed in AOE cells after miR-145 over-expression. In vivo recipients of xenografts of AOE cells that received delivery of miR-145 lentivirus exhibited attenuated tumor formation (Figure 5G).
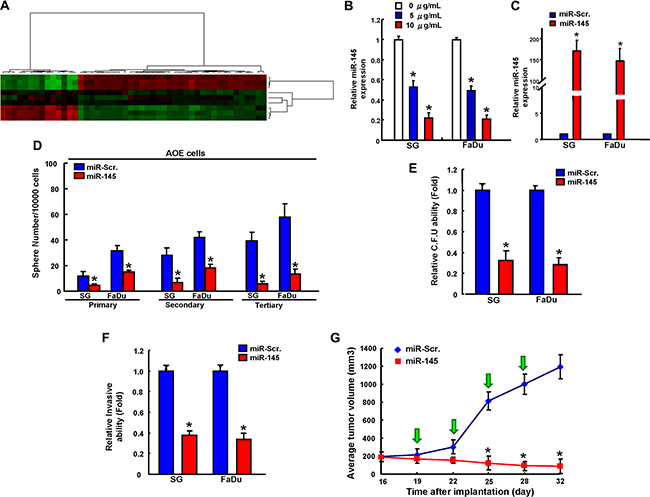
Figure 5: miR-145 effectively reverses long-term arecoline treatment-induced cancer stemness in vitro and in vivo. (A) The heat maps of control and long-term arecoline-treated oral epithelial cells were subjected to miRNA microarray and bioinformatics analysis. A significant decrease in miR-145 expression is found in long-term arecoline-treated oral epithelial cells. qRT-PCR analysis showing miR-145 in long-term arecoline exposed OE cells (B) and theses cells were transfected with miR-145 overexpression vector (C). These miR-145-transfected long-term arecoline exposed OE subclone cells were then assigned for the sphere formation assay (D), colony-forming assay (E), invasion assay (F), and xenografic tumorigenesis in nude mice with IVIS imaging system (G). Data shown here are the mean ± SD of three independent experiments. *p < 0.05 vs. Control.
miR-145 directly targets Oct4 and Sox2 and Oct4/Sox2 co-expression mediates miR-145-regulated cancer stemness and EMT
Subsequently, we illustrate complementarity between the 3′UTR regions of Oct4 and Sox2 and miR-145. To pinpoint the miR-145 target sequences in the 3’UTRs of Oct4 and Sox2, reporter plasmids which contained either full-length or mutated forms of the 3′UTR region of Oct4 and Sox2 were constructed (Figure 6A). Luciferase reporter assays demonstrated that miR-145 reduced the luciferase activity of reporter plasmids containing wild type Oct4 (Figure 6B) and Sox2 (Figure 6C) 3′UTR. However, when the potential Oct4 and Sox2 targeting site was mutated, miR-145 no longer inhibited the luciferase activity (Figure 6B and Figure 6C). Protein levels of Oct4 and Sox2 were also decreased in the miR-145-overexpressing AOE cells (Figure 6D). The functional involvement of Oct4 and Sox2 in miR-145-mediated cancer stemness and EMT in AOE cells was further clarified through Oc4/Sox2 overexpression. miR-145 suppressed spheres-forming capability in AOE cells, which would be rescued by Oc4/Sox2 over-expression (Figure 6E). Furthermore, Oc4/Sox2 over-expression in miR-145-overexpressing AOE cells partially counteracted clonogenicity (Figure 6F) and invasion phenomenons (Figure 6G). We demonstrated that miR-145 down-regulated a pattern of mesenchymal-like transcription factor (Snail and Slug) and up-regulated epithelial protein (E-cadherin) in AOE cells, were reversed by Oct4/Sox2 overexpression (Figure 6H).
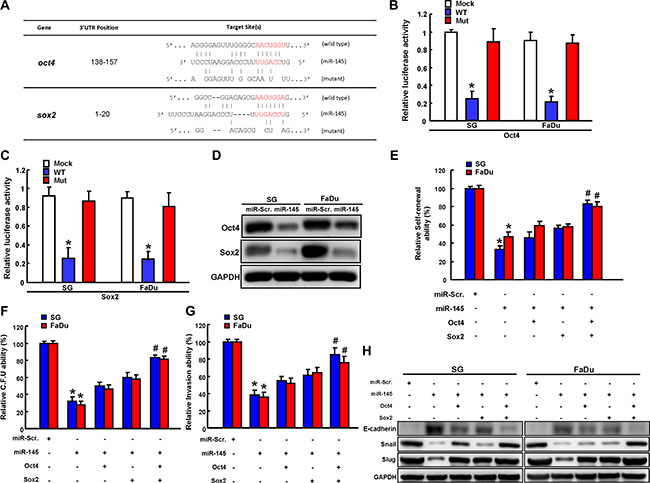
Figure 6: miR-145 regulated by long-term arecoline treatment targets Sox2 and Oct4. (A) Schematic presentation of the constructed Sox2 and Oct4 3′UTR reporter plasmids were used in this study. The wild-type and mutated (Mut) Oct4 (B) and Sox2 (C) reporter plasmids were co-transfected with miR-145 or empty vector. The luciferase activity of each combination was assessed and was presented with wild-type (WT) and mutated (Mut) reporter plasmids. The results of the luciferase assays indicated that only WT reporter activity was inhibited by miR-204. (D) The protein expression levels of Sox2 and Oct4 in transfected indicated plasmids were analyzed by western blot. Long-term arecoline-stimulated oral epithelial cells were transfected with miR145, SOX2, and Oct4 individually or concurrently indicated before being subjected to secondary sphere-forming (E), colony-forming ability assay (F), and invasion assay (G). The percentage or fold-change is presented in the chart. (H) Cells with indicated vectors transfection were then subjected to western blot analysis for the expression level of EMT markers expression.
Negative correlation of miR-145 and Oct4/Sox2 expression in areca nut-associated OSCC patients
To determine significant correlation between miR-145 and Oct4/Sox2, the expression of miR-145 and Oct4/Sox2 among OSCC patients was analyzed by qRT-PCR and Spearman rank correlation tests. qRT-PCR analysis shows a negative correlation between miR-145 expression and Sox2 mRNA expression in OSCC specimen revealed by linear regression analysis (Figure 7A). A significant inverse correlation between miR-145 expression and Oct4 expression was also found using the OSCC tumor samples (Figure 7B). The expression of miR-145 in OSCC tissues was significantly decreased in the recurrent tumor specimens, while miR-145 expression in primary OSCC tissues was higher relative to the recurrent OSCC tissue (Figure 7C). We also found down-regulation of miR-145 in chemo-resistant than in chemo-sensitive OSCC tissues (Figure 7D).
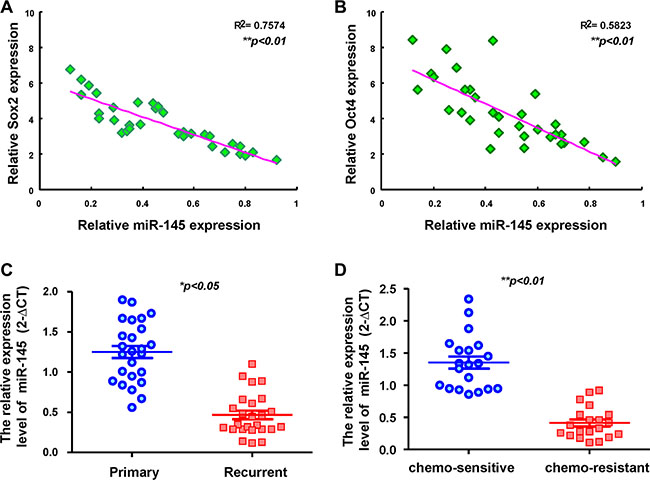
Figure 7: Clinical relevance of miR-145 in OSCC patients. An inverse correlation can be seen between miR-145 /Sox2 (A) and miR-145 /Oct4 (B) in OSCC patients’ tissues. (C) OSCC tissue samples from primary lesions (P) and recurrent (R) lesion were subjected to real-time RT-PCR analysis for the expression levels of miR-145. (D) OSCC tissue samples from chemo-sensitive and chemo-resistant lesions were subjected to real-time RT-PCR analysis for the expression levels of miR-145. **P < 0.01; paired t-test was used in this sample cohort.
DISCUSSION
Epidemiological analysis has demonstrated the majority of OSCC arise in Southeast Asia are due to areca nut chewing together with tobacco smoking [18]. Nevertheless, the pathogenetic roles of areca nut in OSCC have not been well addressed. Epithelial to mesenchymal transition (EMT) process has been implicated as major contributor to oncogenic progression, cancer stemness and cancer metastasis [19]. Previous studies have shown that areca nut extract or arecoline could activate several EMT-related molecules, such as vimentin [20], Snail [21], and ZEB1 [4] in oral epithelial or fibroblastic cells. These interesting findings suggested a possible link between areca nut and EMT/stemness during oral carcinogenesis. In the study, we established chronic arecoline-exposed oral epithelial (AOE) cells. Using our model, these cells exhibit higher capability in oralsphere formation, populations of ADLH-positive cell induction, oncogenicity in vitro and in vivo, and potential for EMT conversion (Figures 1–4). We expect the study is to extend our approaches to insight the influence of arecoline in oncogenic and cancer stemness induction in vitro and in vivo, may allow us to develop potential targeting therapy against OSCC.
A growing number of recent studies have focused on environmental carcinogens exposure leading modulation of tumor malignancy through cancer stemness conversion. Arsenic [22–24] or nickel [25] exposure induced cell transformation through CSCs accumulation. Nicotine or its derivative 4-methylnitrosamino-1-3-pyridyl-1-butanone (NNK) could facilitate CSCs population generation in OSCC [26], breast cancer, colorectal cancer [27], pancreatic cancer [28]. Chronic ethanol exposure can increase the aggressiveness and CSCs of breast cancer cells via p38γ MAPK/RhoC signalings [29]. The areca nut chewing, tobacco, and together with alcohol consumption are the best known etiological factors for the development of OSCC. Future research would be required to examine whether the synergistic effect of these risk factors are involved in CSCs enhancement during OSCC carcinogenesis.
MicroRNAs (miRNAs) are short, non-coding RNAs that regulate gene expression either by translational inhibition or by degradation of the targeted mRNA [30]. It is also well known that dysregulation of miR-145 has been reported in several human solid tumors [31]. miR-145 has been found to be key role in suppressing tumor for OSCC development. miR-145 inhibits cancer stemness in various cancer types, including lung cancer [32], hepatocarcinoma [33], prostate cancer [34], and glioma [35]. This molecule is also associated with the cell cycle [36], differentiation [37], and apoptosis [38], metastasis [39], and chemoresistance [40]. Here, we confirmed that miR-145 negatively regulates cancer stemness in chronic arecoline-exposed OE (AOE) cells. Additionally, we found that Oct4/Sox2 stemness genes are targets of miR-145 through its binding to the Oct4 and Sox2 3′-UTR (Figure 6A). Co-overexpression of Sox2 and Oct4 rescued the repression effect of miR-145 on cancer stemness (Figure 6E–6G). As the Sox2/Oct4 also modulate the EMT of OSCC [41, 42], the regulation of the Sox2/Oct4 by miR-145 that modulate EMT and cancer stemness needs to be further investigated.
In conclusion, chronic arecoline exposure induces malignant phenotype with the acquisition of cancer stemness/EMT, and oncogenicity in vitro and in vivo. miR-145 might be partial mechanism of areca nut-induced OSCC. Validation and detail molecular understanding of later genes are ongoing to determine the arecoline associated pathway alterations during the OSCC tumorigenesis. A full understanding of the mechanism underlying arecoline-induced oral cancer may help to design a more effective strategy to treat areca nut-associated OSCC patients and prolong life.
MATERIALS AND METHODS
Cell culture and reagents
The Smulow–Glickman (S-G) human gingival epithelial and FaDu OSCC cell lines were used in this study. The cultivation of these cells was following the protocols previously used.
Arecoline was purchased from Sigma Chemical Co. (St. Louis, MO¸USA) and was dissolved in ddH2O as a stock solution of 100 mM. Just before use, arecoline was further diluted in culture medium to appropriate final concentrations [41].
Cancer stemness-associated phenotypic analysis
The phenotypes of including oralspheres formation, migration [10], invasion [10], soft agar colony forming assay, ALDEFLUOR assay [9], CD44+ population by flow Cytometry analysis [43] was conducted according to previously used protocols.
Quantitative real-time PCR (qRT-PCR)
Total RNA is prepared from cells using Trizol reagent according to the manufacturer’s protocol (Invitrogen Life Technologies, Carlsbad, CA, USA). qRT–PCRs of mRNAs are reverse-transcribed using the Superscript III first-strand synthesis system for RT–PCR (Invitrogen Life Technologies, Carlsbad, CA, USA). qRT-PCR reactions on resulting cDNAs were performed on an ABI StepOne™ Real-Time PCR Systems (Applied Biosystems).
Western blot analysis
Western blot analysis was followed previously described protocols [4]. The primary antibodies will be those against Oct4, Nanog, Sox2, Snail, Twist, Slug, E-cadherin, and Vimentin [4].
Constructs
miR-145 overexpression plasmid constructs were generated according to our previous methods [9]. The Sox2 and Oct4 3′ UTRs were amplified by PCR and were cloned into the pMIR-REPORT vector (Applied Biosystems). Human full-length Oct4 and Sox2 cDNA was cloned into pCDH1-MCS1-EF1-copGFP (System Biosciences, Cat. No: CD511A-1; Mountain View, CA, USA). Lentivirus production was performed as previously [9].
Xenograft cancer mouse model
All the animal practices in this study has been approved and in accordance with the Institutional Animal Care and Use Committee (IACUC) of Chung Shan Medical University university. Cells from each stable miR-145 -overexpressing or miR-Scramble cells will injected subcutaneously into BALB/c nude mice (6–8 weeks). Tumor volume (TV) will be calculated using the following formula: TV (mm3) = (Length × Width 2) / 2 [6].
OSCC tissue subjects
With the permission of the institutional review board of Chun Shan Medical University, resected tissues from OSCC patients, who gave informed consent for the use of their tissue, were harvested at surgery. OSCC tumor samples were subjected to qRT-PCR analysis.
Statistical analysis
Statistical Package of Social Sciences software (version 13.0) (SPSS, Inc., Chicago, IL) was used for statistical analysis. Student’s t test was used to determine statistical significance of the differences between experimental groups; p values less than 0.05 were considered statistically significant. The level of statistical significance was set at 0.05 for all tests.
ACKNOWLEDGMENTS AND FUNDING
This study was supported by grants from Ministry of Science and Technology (MOST 103-2632-B-040 -001, 104-2632-B-040 -001, 105-2632-B-040 -001) in Taiwan.
CONFLICTS OF INTEREST
The authors have declared no conflicts of interest.
REFERENCES
1. Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer J Clin. 2013; 63:11–30.
2. Li WC, Lee PL, Chou IC, Chang WJ, Lin SC, Chang KW. Molecular and cellular cues of diet-associated oral carcinogenesis—with an emphasis on areca-nut-induced oral cancer development. J Oral Pathol Med. 2015; 44:167–177.
3. Chang MC, Chan CP, Chen YJ, Hsien HC, Chang YC, Yeung SY, Jeng PY, Cheng RH, Hahn LJ, Jeng JH. Areca nut components stimulate ADAM17, IL-1alpha, PGE2 and 8-isoprostane production in oral keratinocyte: role of reactive oxygen species, EGF, JAK signaling. Oncotarget. 2016; 7:16879–94. doi: 10.18632/oncotarget.7621.
4. Ho CM, Hu FW, Lee SS, Shieh TM, Yu CH, Lin SS, Yu CC. ZEB1 as an indicator of tumor recurrence for areca quid chewing-associated oral squamous cell carcinomas. J Oral Pathol Med. 2015; 44:693–8.
5. Lin WT, Shieh TM, Yang LC, Wang TY, Chou MY, Yu CC. Elevated Lin28B expression is correlated with lymph node metastasis in oral squamous cell carcinomas. J Oral Pathol Med. 2015; 44:823–830.
6. Chiou SH, Yu CC, Huang CY, Lin SC, Liu CJ, Tsai TH, Chou SH, Chien CS, Ku HH, Lo JF. Positive correlations of Oct-4 and Nanog in oral cancer stem-like cells and high-grade oral squamous cell carcinoma. Clin Cancer Res. 2008; 14:4085–4095.
7. Prince ME, Sivanandan R, Kaczorowski A, Wolf GT, Kaplan MJ, Dalerba P, Weissman IL, Clarke MF, Ailles LE. Identification of a subpopulation of cells with cancer stem cell properties in head and neck squamous cell carcinoma. Proc Natl Acad Sci USA. 2007; 104:973–978.
8. Chen YS, Wu MJ, Huang CY, Lin SC, Chuang TH, Yu CC, Lo JF. CD133/Src axis mediates tumor initiating property and epithelial-mesenchymal transition of head and neck cancer. PloS one. 2011; 6:e28053.
9. Yu CC, Tsai LL, Wang ML, Yu CH, Lo WL, Chang YC, Chiou GY, Chou MY, Chiou SH. miR145 targets the SOX9/ADAM17 axis to inhibit tumor-initiating cells and IL-6-mediated paracrine effects in head and neck cancer. Cancer Res. 2013; 73:3425–3440.
10. Wu MJ, Jan CI, Tsay YG, Yu YH, Huang CY, Lin SC, Liu CJ, Chen YS, Lo JF, Yu CC. Elimination of head and neck cancer initiating cells through targeting glucose regulated protein78 signaling. Mol Cancer. 2010; 9:283.
11. Yu CC, Hu FW, Yu CH, Chou MY. Targeting CD133 in the enhancement of chemosensitivity in oral squamous cell carcinoma-derived side population cancer stem cells. Head Neck. 2016; 38:E231–8.
12. Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, Campbell LL, Polyak K, Brisken C, et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008; 133:704–715.
13. Lo JF, Yu CC, Chiou SH, Huang CY, Jan CI, Lin SC, Liu CJ, Hu WY, Yu YH. The epithelial-mesenchymal transition mediator S100A4 maintains cancer-initiating cells in head and neck cancers. Cancer Res. 2011; 71:1912–1923.
14. Clay MR, Tabor M, Owen JH, Carey TE, Bradford CR, Wolf GT, Wicha MS, Prince ME. Single-marker identification of head and neck squamous cell carcinoma cancer stem cells with aldehyde dehydrogenase. Head Neck. 2010; 32:1195–1201.
15. Nguyen LV, Vanner R, Dirks P, Eaves CJ. Cancer stem cells: an evolving concept. Nat Rev Cancer. 2012; 12:133–143.
16. Dean M, Fojo T, Bates S. Tumour stem cells and drug resistance. Nat Rev Cancer. 2005; 5:275–284.
17. Ceppi P, Peter ME. MicroRNAs regulate both epithelial-to-mesenchymal transition and cancer stem cells. Oncogene. 2014; 33:269–278.
18. Lu HH, Kao SY, Liu TY, Liu ST, Huang WP, Chang KW, Lin SC. Areca nut extract induced oxidative stress and upregulated hypoxia inducing factor leading to autophagy in oral cancer cells. Autophagy. 2010; 6:725–737.
19. Polyak K, Weinberg RA. Transitions between epithelial and mesenchymal states: acquisition of malignant and stem cell traits. Nat Rev Cancer. 2009; 9:265–273.
20. Chang YC, Tsai CH, Tai KW, Yang SH, Chou MY, Lii CK. Elevated vimentin expression in buccal mucosal fibroblasts by arecoline in vitro as a possible pathogenesis for oral submucous fibrosis. Oral Oncol. 2002; 38:425–430.
21. Lee SS, Tsai CH, Yu CC, Chang YC. Elevated snail expression mediates tumor progression in areca quid chewing-associated oral squamous cell carcinoma via reactive oxygen species. PloS one. 2013; 8:e67985.
22. Sun Y, Tokar EJ, Waalkes MP. Overabundance of putative cancer stem cells in human skin keratinocyte cells malignantly transformed by arsenic. Toxicol Sci. 2012; 125:20–29.
23. Tokar EJ, Qu W, Waalkes MP. Arsenic, stem cells, and the developmental basis of adult cancer. Toxicol Sci. 2011; 120:S192–203.
24. Wong HP, Yu L, Lam EK, Tai EK, Wu WK, Cho CH. Nicotine promotes colon tumor growth and angiogenesis through beta-adrenergic activation. Toxicol Sci. 2007; 97:279–287.
25. Wang L, Fan J, Hitron JA, Son YO, Wise JT, Roy RV, Kim D, Dai J, Pratheeshkumar P, Zhang Z, Shi X. Cancer stem-like cells accumulated in nickel-induced malignant transformation. Toxicol Sci. 2016; 151:376–87.
26. Yu CC, Chang YC. Enhancement of cancer stem-like and epithelial-mesenchymal transdifferentiation property in oral epithelial cells with long-term nicotine exposure: reversal by targeting SNAIL. Toxicol Appl Pharmacol. 2013; 266:459–469.
27. Lee TY, Liu CL, Chang YC, Nieh S, Lin YS, Jao SW, Chen SF, Liu TY. Increased chemoresistance via snail-raf kinase inhibitor protein signaling in colorectal cancer in response to a nicotine derivative. Oncotarget. 2016; 7:23512–20. doi: 10.18632/oncotarget.8049.
28. Al-Wadei MH, Banerjee J, Al-Wadei HA, Schuller HM. Nicotine induces self-renewal of pancreatic cancer stem cells via neurotransmitter-driven activation of sonic hedgehog signalling. Eur J Cancer. 2016; 52:188–196.
29. Xu M, Wang S, Ren Z, Frank JA, Yang XH, Zhang Z, Ke ZJ, Shi X, Luo J. Chronic ethanol exposure enhances the aggressiveness of breast cancer: the role of p38gamma. Oncotarget. 2016; 7:3489–3505. doi: 10.18632/oncotarget.6508.
30. Mendell JT, Olson EN. MicroRNAs in stress signaling and human disease. Cell. 2012; 148:1172–1187.
31. Hu J, Guo H, Li H, Liu Y, Liu J, Chen L, Zhang J, Zhang N. MiR-145 regulates epithelial to mesenchymal transition of breast cancer cells by targeting Oct4. PloS one. 2012; 7:e45965.
32. Hu J, Qiu M, Jiang F, Zhang S, Yang X, Wang J, Xu L, Yin R. MiR-145 regulates cancer stem-like properties and epithelial-to-mesenchymal transition in lung adenocarcinoma-initiating cells. Tumour Biol. 2014; 35:8953–8961.
33. Jia Y, Liu H, Zhuang Q, Xu S, Yang Z, Li J, Lou J, Zhang W. Tumorigenicity of cancer stem-like cells derived from hepatocarcinoma is regulated by microRNA-145. Oncol Rep. 2012; 27:1865–1872.
34. Ozen M, Karatas OF, Gulluoglu S, Bayrak OF, Sevli S, Guzel E, Ekici ID, Caskurlu T, Solak M, Creighton CJ, Ittmann M. Overexpression of miR-145–5p inhibits proliferation of prostate cancer cells and reduces SOX2 expression. Cancer Invest. 2015; 33:251–258.
35. Lee HK, Finniss S, Cazacu S, Bucris E, Ziv-Av A, Xiang C, Bobbitt K, Rempel SA, Hasselbach L, Mikkelsen T, Slavin S, Brodie C. Mesenchymal stem cells deliver synthetic microRNA mimics to glioma cells and glioma stem cells and inhibit their cell migration and self-renewal. Oncotarget. 2013; 4:346–361. doi: 10.18632/oncotarget.868.
36. Xu Z, Zeng X, Xu J, Xu D, Li J, Jin H, Jiang G, Han X, Huang C. Isorhapontigenin suppresses growth of patient-derived glioblastoma spheres through regulating miR-145/SOX2/cyclin D1 axis. Neuro Oncol. 2016;18:830–9.
37. Morgado AL, Rodrigues CM, Sola S. MicroRNA-145 Regulates Neural Stem Cell Differentiation Through the Sox2-Lin28/let-7 Signaling Pathway. Stem Cells. 2016. 34:1386–95.
38. Gomes SE, Simoes AE, Pereira DM, Castro RE, Rodrigues CM, Borralho PM. miR-143 or miR-145 overexpression increases cetuximab-mediated antibody-dependent cellular cytotoxicity in human colon cancer cells. Oncotarget. 2016; 7:9368–9387. doi: 10.18632/oncotarget.7010.
39. Li YQ, He QM, Ren XY, Tang XR, Xu YF, Wen X, Yang XJ, Ma J, Liu N. MiR-145 inhibits metastasis by targeting fascin actin-bundling protein 1 in nasopharyngeal carcinoma. PloS one. 2015; 10:e0122228.
40. Zhu X, Li Y, Xie C, Yin X, Liu Y, Cao Y, Fang Y, Lin X, Xu Y, Xu W, Shen H, Wen J. miR-145 sensitizes ovarian cancer cells to paclitaxel by targeting Sp1 and Cdk6. Int J Cancer. 2014; 135:1286–1296.
41. Tsai LL, Hu FW, Lee SS, Yu CH, Yu CC, Chang YC. Oct4 mediates tumor initiating properties in oral squamous cell carcinomas through the regulation of epithelial-mesenchymal transition. PloS one. 2014; 9:e87207.
42. Chou MY, Hu FW, Yu CH, Yu CC. Sox2 expression involvement in the oncogenicity and radiochemoresistance of oral cancer stem cells. Oral Oncol. 2015; 51:31–39.
43. Hu FW, Tsai LL, Yu CH, Chen PN, Chou MY, Yu CC. Impairment of tumor-initiating stem-like property and reversal of epithelial-mesenchymal transdifferentiation in head and neck cancer by resveratrol treatment. Mol Nutr Food Res. 2012; 56:1247–1258.