
Cancer stem cells (CSCs)
Cancer stem cells (CSCs) are believed to be a small subpopulation of tumour cells that have properties of tumorigenesis, multilineage differentiation potential, self-renewal [1], slow cycling capacity [2] and tumorigenicity [3, 4]. In recent years, methods enabling tumorigenic cells and their progeny to be tracked and clearly observed in vivo have been developed, making the existence of CSCs increasingly more convincing [5-7].
CSCs are at a less-differentiated state than corresponding cancer cells. Similar to other stem cells, CSCs possess the capacity for asymmetrical division in addition to symmetrical division [8-10]. During asymmetrical division, CSCs divide into two different daughter cells, one of which copies the mother cell’s entire genome, while the other has fewer features of stemness. Due to their ability to divide asymmetrically, CSCs possess the capacity for self-renewal and tumour initiation [10]. These properties of asymmetrical division and self-renewal enable CSCs to maintain dynamic control of their numbers, and tumours invariably consist of a mixture of CSCs and their diversely differentiated progeny, contributing to the significant phenotypic and functional heterogeneity of CSCs [11]. Due to their self-renewal and tumour-initiating properties, CSCs are believed to be the starting point for cancer and are thought to play key roles in cancer relapse and metastasis [12, 13]. As a result, CSCs have become a promising target for preventing cancer relapse and for vastly improving the survival of cancer patients [14-16].
CSCs are often dormant and remain in the CSC niche, which protect them from damage by any of the existing anti-tumour therapies [14, 17-19]. The CSC niche is a favourable environment for CSCs to achieve an optimal balance between self-renewal, activation and differentiation [20, 21]. In response to stress, CSCs are able to be “activated” and recruited into other tissues, where they differentiate and generate malignant cells [19]. Blagosklonny, M.V. noted that quiescent CSCs play a negligible role in advanced cancers that have a poor response to therapy and that only “activated” CSCs contribute to proliferation, progression and therapeutic failures. As such these cells should be targeted and eliminated [22, 23]. However, Gupta, G.B. and colleagues have discovered that cancer cells in various states were able to stochastically transit between states and generate a phenotypic equilibrium in breast cancer [24], indicating that immortal, quiescent CSCs, and even non-CSCs could be able to transit into proliferating CSCs when proliferating CSCs are eliminated [25-28].
Circulating tumour cells (CTCs), which are present in the blood, and disseminated tumour cells (DTCs), which are located in a secondary organ, are positively associated with tumour metastasis, relapse and poor survival [29-33]. Interestingly, CTCs and DTCs display the phenotypes of both CSCs and epithelial-mesenchymal transition (EMT) [34-37]. It is hypothesized that these CTCs and DTCs can evade immune targeting by undergoing EMT and losing their epithelial-related features. In this way, they achieve a more “de-differentiated” status and maintain more features of stemness while retaining their malignancy [33, 38]. In breast cancer, the proportion of CSCs in primary cancer is supposed to be less than 1% [39], whereas approximately over 50% of CTCs express EMT and CSC markers [40]. However, the relationship between CTCs, DTCs and CSCs is complicated and remains a topic of debate.
Cancer is known to be a heterogeneous disease [41-43]. First, there is inter-tumour heterogeneity, which involves different degrees of aggressiveness and clinical outcomes between patients who have the same tumour type. Second, there is intra-tumour heterogeneity, which involves biological and molecular differences between the tumour cells within the same tumour in a single patient [41, 44]. Cancer heterogeneity may be associated with the CSC content [45]. Histologically, tumours with a high percentage of CSCs may be poorly differentiated, undifferentiated or mixed tumours. As proposed by Weinberg RA et al., tumours are a heterogeneous mixture of CSCs that have mixed epithelial-mesenchymal phenotypes and non-stem cells that are epithelial [46]. By undergoing EMT, epithelial cells may acquire stem-like features [47-49]. However, only non-stem epithelial malignancies are attenuated or eliminated by existing anticancer treatments; by contrast, CSCs escape and survive [50]. As a result, to eliminate the cancer root, CSCs need to be specifically targeted and eliminated.
Multiple stem-cell specific markers and functional assays have been used to identify putative CSCs for both in vitro and in vivo studies. Some CSC-related markers are being developed as potential cancer therapy targets. However, it remains challenging to target CSCs because of their complex biology and instability [46, 51, 52]. A related problem is that the frequency and identity of tumorigenic cells varies from patient to patient, indicating that CSC markers identified in one tumour may not be sufficient for identifying CSCs in another tumour [53]. To date, no universal CSC marker for identifying CSCs in all tumours has been identified. Furthermore, CSCs reside in a CSC niche, which may preserve their phenotypic plasticity, protect them from the immune system and maintain their dynamic number and state [53-56], thereby making it more difficult to target them.
Cervical cancer stem cells (CCSCs)
Cervical carcinoma is the most common type of gynaecological malignancy worldwide [57], and it is one of the leading causes of cancer death among females, especially in less developed countries [58]. In contrast to the traditional “clonal evolution” theory of carcinogenesis, which describes cervical carcinoma as a consequence of unlimited and uncontrolled cellular proliferation, there is heterogeneity in cervical carcinoma. Intra-tumour genetic heterogeneity in cervical carcinoma is associated with a poor chemo/radio-therapy response [59], lymph node metastasis and pelvic recurrence [60]. One explanation for the heterogeneity in cervical carcinoma is the existence of CCSCs. Because of the asymmetrical division of CCSCs, cervical carcinoma tissue consists of diversely differentiated carcinoma cells. Furthermore, CSCs have recently been found to be capable of transdifferentiation into vascular endothelial cells and other tumour-associated stromal cells [61], which may also contribute to tumour heterogeneity.
Cervical carcinoma is known to have a causal relationship with specific human papillomavirus (HPV) strains [62]. However, not all cervical epithelial cells infected with carcinogenic HPV will generate cervical carcinoma. After infection with carcinogenic HPV, cells located in the transition area between the endocervix and exocervix, which is known as the squamo-columnar (SC) junction, more easily lead to cervical intraepithelial neoplasia (CIN) and carcinogenesis [63, 64]. Studies have shown that approximately 90% of CIN3 and cervical cancer arise within or very near the SC junction [64]. These specific SC junction cells exhibit unique morphology and gene profiles, which distinguish them from the adjacent endocervical and ectocervical epithelium. For instance, they express SC junction-specific markers, keratin 7 (Krt7), anterior gradient 2 (AGR2), cluster differentiation 63 (CD63), matrix metalloproteinase 7 (MMP7) and guanine deaminase (GDA). Intriguingly, these junction-specific markers are also expressed in carcinogenic HPV-associated CINs and carcinomas, including both squamous cell carcinomas and adenocarcinomas, indicating that multiple cervix malignancy subtypes are derived from the SC junction cells [63, 65]. Similarly, ovarian carcinoma originates in the transition area between the ovarian surface epithelium, mesothelium and tubal epithelium, and these junction cells have long-term stem cell properties ex vivo and in vivo, as determined by serial sphere generation and long-term lineage-tracing assays. As a result, the junction area has been considered to be an important cancer stem cell niche in which ovarian cancer originates [18]. We hypothesize that cervical carcinoma develops from stem-like cells in the transition area of the cervical opening that are infected with carcinogenic HPV and that the junction/transition area may be a possible niche for CCSCs (Figure 1).
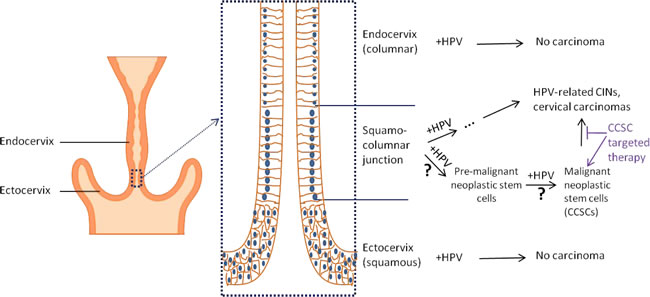
Figure 1: Illustration of cervical carcinogenesis and CCSCs. Carcinogenic HPV infection has a causal relation with cervical carcinogenesis. However, when the cervix is infected with carcinogenic HPV, HPV-related CINs and cervical carcinomas are usually generated within a specific cell population that is located in the ectoendocervical squamocolumnar (SC) junction of the cervix. They are typically not generated in the columnar cells located within the endocervix and squamous cells within the ectocervix. The HPV-related CINs and cervical cancers maintain the genetic profile of the junction cells, indicating their cellular hierarchy. Progenitor cells located in the junction area infected with carcinogenic HPV are likely to become pre-malignant neoplastic stem cells that can propagate malignant neoplastic stem cells (CCSCs), which propagate cervical carcinoma clones. Therapeutically targeting these cells may prevent the propagation of HPV-related CINs and cervical carcinomas.
Cervical cancer treatment includes surgery, chemotherapy and radiation therapy, and these approaches have improved carcinoma survival. Cisplatin-based chemotherapy is a commonly used cervical cancer therapy [66]. However, some CCSCs that were identified by multi-marker and functional examinations are resistant to cisplatin-based chemotherapy [67] and radiation therapy [68], which is in agreement with the chemo/radio-resistance that is generally observed with CSCs.
Considering the tumorigenic potency of common CSCs, CCSCs may be the route of cervical carcinogenesis and lead to distant metastasis. According to this theory, therapeutic treatment that specifically targets CCSCs may prevent the generation of new tumours. Such a treatment approach would offer a critical tool for preventing distant metastasis, tumour relapse and chemo/radio-resistance. With the combination of CCSC-targeted therapy and traditional anti-cancer therapy, cervical cancer might someday be curable. To achieve this goal, the first requisite and most challenging problem is how to specifically identify and target CCSCs as part of cancer treatment.
However, just as CSCs have a CSC niche, CCSCs may also reside in their own niche, which may differ from the tumour microenvironment. The CSC niche is not well understood, and the characteristics, cellular communication and molecular profile of the CCSC niche are even more obscure, making the identification of CCSCs even more challenging. The existing approaches for identifying CCSCs primarily depend on the expression of specific stem cell markers in malignant cells and their stem-like functional properties.
Stem cell markers for CCSCs
Stem cell markers are widely used to identify CSCs for both in vitro and (especially) in vivo studies because of the practical limitations of functional assays. Therefore, targeting CSC-specific markers is one of the most promising and easily achievable approaches for identifying CSCs, even though molecular assays are not sufficient to fully define CSCs [69]. Stem cell markers for CCSCs have been accumulating, and novel markers are being identified. We reviewed and summarized the markers for CCSCs that are currently used or are potential candidates in CCSC studies (Table 1). However, as with the heterogeneity of CSCs, CCSC markers vary from tumour to tumour. Consequently, cervical cancer cells that express a single stem cell marker do not always qualify as CCSCs.
Table 1: CCSC markers that are currently in use
marker |
Cell lines |
Establishment of stem-like cells |
Stem-like properties |
Clinical associations |
ABCG2 |
SiHa, CaLo, C-33A |
SP |
High colony forming efficiency, multilineage differentiation, asymmetrical division [78] |
|
ALDH1 |
ALDH1+ cells by FACS using Aldefluor staining |
High cell proliferation, migration, sphere forming efficiency [84], limiting dilution tumorigenicity, multilineage differentiation, asymmetrical division, self-renewal, highly expressing OCT4, NANOG, KLF4 and BMI1 [67] |
Poor survival [83] |
|
CD133 |
Hela |
SP |
High cell proliferation, self-renewal, chemo/radio-resistance, limiting dilution tumorigenicity, multilineage differentiation, anti-apoptosis, highly expressing OCT4, ABCG2, SOX2 [98-100] |
|
CD49f |
HeLa, SiHa, Ca Ski, C-4 I |
Tumorigenic spheroids |
Limiting dilution tumorigenicity, self renewal, highly expressing stem cell markers and EMT markers, radioresistance [68] |
|
OCT4 |
HeLa, SiHa |
OCT4 over-expressing cells by plasmid transfection |
Enhanced tumorigenicity, anti-apoptosis [113] |
Poor differentiation [111], lymph node metastasis [111, 113], radioresistance, poor survival [112] |
OPN |
HeLa, SiHa |
OPN over-expressing cells by plasmid transfection |
Enhanced tumor growth, stimulating CD44 phosphorylation and CD44-dependent MAPK and NF-κB activation [123] |
Hypoxic radiation resistance and poor survival [121] |
SOX2 |
SiHa, C33A |
SOX2 over-expressing cells by plasmid transfection and cell sorting by SOX2 antibody by FACS |
Differentiation, self-renewal, enhanced tumorigenicity, highly expressing stem cell markers OCT4, ALDH1, BMI1 and EMT-related markers vimentin, snail, β-catenin [134] |
Higher SOX2 expression in cervical carcinoma than normal cervix [129-131], in high grade of dysplasia than low grade of dysplasia [133], enhanced radioresistance, poor survival [112] |
ABCG2
ATP-binding cassette sub-family G member 2 (ABCG2), also known as breast cancer resistance protein (BCRP), is a drug efflux membrane transporter of the ATP-binding cassette (ABC) family. ABCG2 pumps out a wide variety of chemical compounds from cells and plays a major role in multi-drug resistance in a number of cancer types [70]. It is also a determining molecular marker in a side population (SP) phenotype, which is considered to be a characteristic feature of CSCs [71, 72]. Therapies that target and block the function of ABCG2, such as Axitinib [73] and Icotinib [74], could enhance cellular sensitivity to chemotherapy. ABCG2 has thus become a CSC marker and a potential target for cancer treatment [75]. The expression and activity of ABCG2 may interact with various lipid compounds, especially those residing in close proximity within the plasma membrane [76]. In cervical cancer, the redox sensing factor Nrf2 may play an important role in the transcriptional regulation of ABCG2, and cells with upregulated Nrf2 and ABCG2 exhibit stem-like characters, including infinite cell proliferation, longevity and prevention of apoptosis [77]. Three commercial cell lines, SiHa, CaLo, and C-33A, were sorted using fluorescence-activated cell sorting (FACS) to obtain SP and non-SP (NSP) cells. The SP cells had high ABCG2 expression and colony forming efficiency, as well as the capacity to generate both SP and NSP cells, suggesting that ABCG2 plays a pivotal role in maintaining cell stemness [78].
ALDH1
Aldehyde dehydrogenase 1 (ALDH1) is a metabolic enzyme that is localized in the cytoplasm and catalyses the dehydrogenation of aldehydes. ALDH1 is also associated with the tumorigenic cell fraction, capacity for self-renewal and tumorigenesis [79], especially in breast cancer [80, 81]. Additionally, the presence of ALDH1-positive CSC-like cells in primary breast cancer is associated with the successful establishment of patient-derived xenografts [82].
ALDH1 expression in cervical carcinoma patient tissues is an independent risk factor that indicates a poorer survival probability [83]. High ALDH1 expression in cervical cancer cells is associated with a high rate of cell proliferation, sphere formation, migration and tumorigenesis [84], indicating that it acts as a stemness factor in cervical cancer. To determine whether ALDH could serve as a specific marker of CCSCs, ALDH-high and ALDH-low cells were sorted from 4 cervical cancer cell lines and 5 primary tumour xenografts using FACS and were then examined for the presence of CSC characteristics. ALDH-high cells had a higher tumorigenicity potential in vivo than ALDH-low cells, and they could divide into both ALDH high and low cells in vitro and in vivo, thereby establishing a cellular hierarchy and enhancing self-renewal and differentiation potentials [67].
CD133
CD133 is a pentaspan transmembrane glycoprotein (120 kDa) encoded by the prominin 1 (PROM1) gene in humans [85]. CD133 has been extensively used as a CSC marker in many tumours [86], including brain [87], breast [88], colon [89], liver [90, 91], lung [92], melanoma [93, 94] and ovarian [95-97] cancers, although there are limited reports on the relationship between CD133-positivity and the stem-like characteristics of tumour cells. In cervical cancer cells, it was found that the SP cells that were sorted using FACS from the HeLa line, which displays the stem-like probabilities of proliferation, differentiation, self-renewal, chemo/radio-resistance and tumorigenicity, had high CD133 expression compared with the NSP cells [98, 99]. As a result, CD133 may serve as a specific CCSC marker in CSC-targeted therapy. Other studies have also shown that established cervical stem-like cells express CD133 and other CSC-related markers and exhibit radiation resistance [68, 100].
CD49f
CD49f is a cell surface protein that is encoded by the integrin alpha 6 (ITGA6) gene. It is highly expressed in human embryonic stem cells (ESCs) [101], mesenchymal stem cells (MSCs) [101, 102] and haematopoietic stem cells (HSCs). In MSCs, CD49f enhances multipotency and maintains stemness by directly regulating thetranscription factors POU class 5 homeobox 1 (POU5F1, OCT4) and SRY-box2 (SOX2) [101]. HSCs can be purified from mobilized peripheral blood cells based on CD49f expression [103]. CD49f and CD44, a stem cell marker, are transcriptionally upregulated by Y-box binding protein-1 (YB-1) in breast cancer cells, enhancing the stem-like properties of these cells, which include self-renewal, colony forming efficiency and drug resistance [104]. CD49f is also involved in maintaining stem-like features in multiple cancer types, including gastric, colon, and prostate cancer [105-108]. Furthermore, CCSC models have been established in cell line-developed tumorigenic spheroids that are resistant to radiotherapy, and these CCSC models have high CD49f expression [68].
OCT4
OCT4, also known as OCT3 and OCT3/4, is a transcription factor that is expressed by the POU class 5 homeobox 1 (POU5F1) gene in humans. It plays a key role in embryonic development and the maintenance of stem cell pluripotency [109, 110]. OCT4 is over-expressed in cervical cancer tissues compared to adjacent normal tissues [111], and the over-expression of OCT4 in cervical cancer cells is associated with a low-differentiation grade of cervical cancer cells and positive lymph node metastasis [111]. Clinical data have shown that high OCT expression is positively associated with radiotherapy resistance, and OCT expression has been found to be an independent risk factor for cervical cancer patient survival [111, 112]. Furthermore, an in vitro study demonstrated that OCT4 promotes tumorigenesis and inhibits cancer cell apoptosis [113].
At least three isoforms (OCT4A, OCT4B and OCT4B1), which are produced by alternative splicing, were discovered, and they may play different roles in stem cell biology [114, 115]. Among the three identified isoforms, nuclear OCT4A is the most widely studied and has been recognized as a key factor in regulating pluripotency [115, 116]. In cervical carcinoma, both nuclear OCT4A and cytoplasmic OCT4B have been reported to be over-expressed [117]. OCT4A is responsible for the maintenance and self-renewal of CCSCs, and it may serve as a CCSC marker, while cytoplasmic OCT4B may work with OCT4A to regulate cervical carcinoma progression by inducing angiogenesis and EMT [117].
Osteopontin (OPN)
Osteopontin (OPN), a chemokine-like extracellular matrix protein, is secreted by malignant cells and tumour stromal cells. OPN is a key mediator of tumour cell migration and metastasis [118, 119]. OPN was primarily found to be an endogenous hypoxic marker because it is upregulated by hypoxia [120] and tends to bind to the hypoxic regions of tumour tissues. The over-expression of OPN predicts hypoxic radiation resistance and poor survival in human cervical cancer [121]. OPN may also induce tumour angiogenesis by modulating HIF1α-dependent VEGF expression in response to hypoxia [122]. OPN over-expression in a murine xenograft model of human cervical cancer enhanced tumour growth; conversely, OPN silencing, mediated by short hairpin RNA, blocked this effect [123]. OPN could also be detected in the blood, and an elevated serum OPN level predicts poor survival in cervical cancer patients [124].
OPN expression in hepatocellular carcinoma cell lines is associated with high SP fractions, spheroid formation and tumorigenicity rates in immunodeficient mice [125]. OPN can bind to CD44 receptor family members and regulate tumour cell fate through OPN-CD44 signalling [125]. OPN regulates CD44-mediated p38 phosphorylation, affecting downstream genes, nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and NF-κB-dependent expression of furin, which are involved in the response of human papilloma virus (HPV) [123]. OPN-CD44 signalling may enhance cancer stem-like features [126], and the OPN-mediated self-renewal capabilities may be suppressed by the reduction in NF-κB expression [125].
SOX2
SOX2 is a key transcription factor that is involved in embryonic development and plays a critical role in determining stem cell fate [127, 128]. There is significantly higher nuclear SOX2 expression in cervical carcinoma than in normal cervix tissues [129-131]. Furthermore, cervical cancer with high SOX2 expression is more poorly differentiated [130], indicating that SOX2 might be a marker for undifferentiated cervical cancer. Both in vitro and in vivo studies have shown that cervical cancer cells with SOX2 over-expression have increased cell proliferation [132], clonogenicity, and tumourigenicity [129]. Oncogenic virus HPV-positive cervix cells that generate a higher grade of dysplasia often have higher SOX2 expression than those that generate a lower grade of dysplasia [133]. This indicates that SOX2 plays an important role in early tumour initiation. Moreover, cervical squamous cancer patients with high SOX2 expression in tumour cells have enhanced radiation resistance [112]. The cervical cancer cell lines, SiHa and C33A, were transfected with a plasmid containing the human SOX2 gene, and were sorted using a SOX2 antibody and FACS. The SOX2-positive population expressed higher levels of stem cell-related genes, OCT4 and ALDH1, and EMT-related genes, indicating that CCSCs were more likely to be SOX2 positive cells [134]. Moreover, the SOX2 positive population showed higher probabilities for differentiation, self-renewal and tumourigenicity [134], which further confirms the role of SOX2 as a practical CCSC marker in stem-cell functional examinations.
Other potential markers for CCSCs
There are a number of specific markers for CSCs that could potentially serve as CCSC markers, although little evidence supports their use for this purpose. These include the BMI1 proto-oncogene, polycomb ring finger (BMI1) and Kruppel-like factor 4 (KLF4). BMI1 is a core transcription factor involved in the regulation of EMT and CSC self-renewal, and it integrates multiple signalling pathways, such as TWIST1 [135, 136]. KLF4, as a well-known member of the KLF family, is one of the four factors that can reprogram adult fibroblasts into induced pluripotent stem cells. The significance of KLF4 in CSC regulation has recently increased [137-139]. BMI1 and KLF4 expression levels were found to be elevated in ALDH-high cervical cancer cells that display high stem-like features compared to the ALDH-low cell population [67], indicating that BMI1 and KLF4 may be CCSC markers.
CD44
CD44 has been accepted as a CSC marker in a number of tumours, including gastric cancer [140], colorectal cancer [141-143], glioma [126], head and neck cancer [144, 145] and breast cancer [146]. Many functionally distinct isoforms could be encoded by the CD44 gene because of the complex alternative splicing of transcripts, and these isoforms may participate in different oncogenic signalling pathways and play different roles in tumour progression [147, 148]. Among these splicing variants, the smallest isoform, CD44s, which lacks all variant exons, is the standard CD44 [149] and the one associated with CSCs [148, 150, 151]. However, an immunohistochemical (IHC) study demonstrated that CD44s could be largely expressed in both normal cervix and cervical cancer, and soluble CD44s in serum was also found in both normal and invasive cervix [152]. Sorted SP and NSP cells from the HeLa line, which showed significant differences in many stem-like characteristics, did not have significant differences in their CD44 expression [99]. To date, the evidence for CD44 as a specific CCSC marker remains insufficient, although it is widely used as a general CSC marker in many tumours.
C-Kit
A proto-oncogene, c-Kit, which is also known as tyrosine-protein kinase KIT or CD117, is a transmembrane cytokine receptor expressed on the surface of haematopoietic stem cells and other cell types. It is normally phosphorylated and activated by binding to the KIT ligand, which is also called stem cell factor [153]. It is a widely used stemness marker for recognizing cancer stem cells in various tumour types, including ovarian cancer [154-156], endometrial cancer and osteosarcoma [155, 157, 158]. In HPV-associated cervical cancer, SCF-activated c-Kit may activate interleukin-2 receptor betagamma signalling in the absence of IL-2, promoting T cell proliferation [159]. Unusual expression of c-Kit was found in cervical squamous cell carcinoma without c-Kit gene amplification [160], and the tumours with this unusual c-Kit expression had a high DNA methylation ratio and hypermethylation of the c-KIT promoter [161], suggesting that the c-Kit expression may be regulated by DNA methylation. However, the evidence that c-Kit serves as a stem cell marker for cervical cancer is not yet comprehensive.
NANOG
NANOG homeobox (NANOG) is another transcription factor, in addition to SOX2 and OCT4, which is highly expressed in human ESCs. NANOG plays an essential role in ESC maintenance of pluripotency and the regulation of proliferation and asymmetric division [162, 163]. NANOG has been broadly reported as a CSC marker that regulates self-renewal and tumorigenesis in many tumour types [102, 164-172]. The human form of NANOG has 11 pseudogenes, of which NANOGP8 encodes a functional protein product with only 3 amino acids that differ from the product of NANOG [173]. An IHC study clarified the expression of NANOG in patients with cervical cancer and cervical dysplasia, and its expression was found to be significantly higher in cervical cancer than in cervical dysplasia and higher in cervical dysplasia than in normal cervical epithelia [174], supporting the role of NANOG in carcinogenesis and cervical carcinoma progression. Interestingly, according to the IHC study, in cervical cancer cells and surrounding stromal cells, NANOG was frequently observed in the cytoplasm, instead of the nucleus, where it is found in many other cell types [175]. The cytoplasmic NANOG that is expressed in stromal cells may promote cervical cancer progression [175].
Functional assays for identify-ing CCSCs
As in normal stem cells, functional assays based on stemness-specific properties are the gold standard criteria for identifying CSCs. Functional verification of CCSCs is based on the same rules that are used for CSCs. They include clonogenic activity in soft agar, sphere-forming efficiency in non-adherent cultures, examination of self-renewal and the differentiation potential, and tumorigenic capacity according to the limiting dilution tumorigenicity assay [69]. Asymmetrical cell division is a characteristic and marked manifestation of self-renewal and is assayed by checking whether the “purified” test cells can divide into two populations, one that mimics the parent cell and another with different or more differentiated features. In some cases, the presence of an activated self-renewal pathway is thought to be typical, e.g., PTEN/Akt/ β-catenin signalling and PTEN/Akt/PI3K signalling [176, 177]. Anti-apoptosis may also be a discriminating characteristic of CSCs [178]. The presence of chemo-resistance and radiation resistance also helps characterize CSCs [179, 180]. Nevertheless, all CSC-specific functional assays have been performed in vitro, and it remains challenging to examine the CSC-related functions of potential CSCs in vivo. Therefore, the role of CSC markers in identifying CSCs in vivo is currently emphasized, although they are not the standard criteria and are insufficient for specifically identifying CSCs.
SP is widely recommended as another test for identifying CSC populations [181-185]. The SP discrimination assay is a method that uses flow cytometry to detect stem cells and CSCs based on the dye efflux properties of the ABC family of transporter proteins expressed within the cell membrane [186]. The sorting method based on SP has been considered to be simple and effective in cancer stem cell research [187]. A human cervical cell line, HeLa, was sorted using FACS into SP and NSP cells, and the SP population had higher expression of a CCSC marker, CD133, and displayed most of the classic CSC characteristics, such as increased proliferation, self-renewal, differentiation potential, tumorigenicity and chemo/radio-resistance [98, 99]. However, the SP phenotype is not exclusive to stem cells and is not universal in all cancer types [188]. The procedure for SP population detection is continually being optimized to achieve more specific and sensitive results [186].
NOVEL STRATEGIES AND CHALLEN-GES FOR CSC TARGETED THERAPY
To improve the current treatment of cancer and prevent cancer relapse, CSC-targeted therapy has been intensively studied in recent years. However, to date, studies on specific CCSC targeted therapies are very limited. We will generally discuss CSC targeted therapies, which may hint at potential strategies for CCSC targeted therapies. CSC-specific markers and signalling pathways have been largely used as therapeutic targets, and a variety of CSC targeting strategies are being studied.
The dual-targeting strategy has been proposed to target CSCs in recent years [189]. VS-5584, as a potent and selective dual inhibitor of mTORC1/2 and class I PI 3-kinases (PI3K), specifically targets human CSCs and inhibits their tumour initiating capacity, as demonstrated in murine xenograft models with human breast and ovarian cancers [190]. More studies on dual-targeting therapy that could be used to target CSCs are under investigation. For instance, at the American Association of Cancer Research (AACR) 106th Annual Meeting 2015, dual targeting of delta-like ligand 4 (DLL4) and programmed death 1 (PD1) was demonstrated to be a promising cancer therapy [191]. However, there is insufficient information to make a conclusive statement about the therapeutic potential of the dual-targeting strategy, and no studies have yet reported on the use of dual-targeting to treat CCSCs. More studies on dual-targeting are warranted.
CSC targeting with nanoparticles (NPs) is another novel, possibly effective therapeutic approach that has been a topic of recent, exciting investigations [192-195]. NP-enabled therapies have been designed to inhibit stem cell-related functions by targeting stem cell-specific signalling pathways (e.g., Wnt/β-catenin [196], Notch [197] and reactive oxygen species (ROS) signalling [198]) and/or CSC-specific markers (e.g., CD44 [199, 200]) that are critically involved in maintenance of cell stemness [201]. Kumar P et al. proposed an approach for targeting endothelial cells in the cancer stem cell niche using a twin NP of iron coated with gold [202]. Gold NPs that were further conjugated with sophorolipids were found to be effective in the treatment of human glioma stem cells [203]. NP-mediated photothermal therapy is effective for both breast cancer stem cells and non-stem cancer cells [204]. A hybrid NP of bioactive quinacrine and silver was also shown to enhance cytotoxicity and inhibit oral stem cells in vitro [205]. However, NP-enabled therapies remain far from an ideal CSC-specific targeting therapy, especially because sensitive, specific markers or an equal combination of different markers and distinctive CSC signalling pathways have not yet been characterized for each tumour type.
Although studies on CSC-targeting therapies have increased in recent years, there remain some limitations that are not easy to overcome. CSCs are typically present at very low levels in tumours, accounting for only approximately 0.1-10% of tumour cells [206]. Furthermore, most of the currently available information on CSC targeting therapy is largely inspired and influenced by the biological characteristics of normal stem/progenitor cells, such as their discriminating surface markers and specific signalling pathways. As a result, CSC-targeted therapy may damage normal stem/progenitor cells and block the regeneration of normal tissues, causing tissue or organ dysfunction.
Conclusions
Cancer is a heterogeneous disease that consists of a small subpopulation of CSCs with mixed epithelial-mesenchymal phenotypes and non-stem cells with an epithelial phenotype. CSCs have become promising targets for cancer treatment due to their capacity for self-renewal and tumorigenicity. CSC-specific markers and functional assays are widely used to identify CSCs. However, because of their practical limitations, functional examinations for CSCs are not as commonly used as CSC-specific markers in in vivo studies.
CCSCs, which can generate multiple tumour cells, seed distant metastases and promote tumour recurrence, are becoming promising targets for treating cervical carcinogenesis. HPV-associated cervical carcinoma may arise from the HPV-infected cells in the SC junction area, which may act as the CCSC niche. Several markers, such as ABCG2, ALDH1, CD133, CD49f and SOX2, were identified as CCSC-specific markers in well-known cervical cancer cell lines based on their stem-like functional probabilities in vitro. The targeting of various stem cell related-markers and signalling pathways may offer a novel strategy for CSC-targeted therapy, such as through dual-targeting and NP-enabled therapies. Challenges for CSC targeted therapy remain, including potential damage to normal stem/progenitor cells.
Acknowledgments
Financial support was received from the Norwegian Cancer Society (71409 – PR-2006-0203) and the South-Eastern Norway Regional Health Authority (2015035).
Conflicts of interest
The authors declare no conflict of interest.
References
1. Kreso A and Dick JE. Evolution of the cancer stem cell model. Cell Stem Cell. 2014; 14:275-291.
2. Velasco-Velazquez MA, Yu Z, Jiao X and Pestell RG. Cancer stem cells and the cell cycle: targeting the drive behind breast cancer. Expert Review of Anticancer Therapy. 2009; 9:275-279.
3. Fitzgerald TL and McCubrey JA. Pancreatic cancer stem cells: Association with cell surface markers, prognosis, resistance, metastasis and treatment. Advances in Biological Regulation. 2014; 56:45-50.
4. Yang L, Ren Y, Yu X, Qian F, Bian BS, Xiao HL, Wang WG, Xu SL, Yang J, Cui W, Liu Q, Wang Z, Guo W, et al. ALDH1A1 defines invasive cancer stem-like cells and predicts poor prognosis in patients with esophageal squamous cell carcinoma. Modern Pathology. 2014; 27:775-783.
5. Driessens G, Beck B, Caauwe A, Simons BD and Blanpain C. Defining the mode of tumour growth by clonal analysis. Nature. 2012; 488:527-530.
6. Chen J, Li Y, Yu TS, McKay RM, Burns DK, Kernie SG and Parada LF. A restricted cell population propagates glioblastoma growth after chemotherapy. Nature. 2012; 488:522-526.
7. Schepers AG, Snippert HJ, Stange DE, van den Born M, van Es JH, van de Wetering M and Clevers H. Lineage tracing reveals Lgr5+ stem cell activity in mouse intestinal adenomas. Science. 2012; 337:730-735.
8. Beier D, Hau P, Proescholdt M, Lohmeier A, Wischhusen J, Oefner PJ, Aigner L, Brawanski A, Bogdahn U and Beier CP. CD133(+) and CD133(-) glioblastoma-derived cancer stem cells show differential growth characteristics and molecular profiles. Cancer research. 2007; 67:4010-4015.
9. Locke M, Heywood M, Fawell S and Mackenzie IC. Retention of intrinsic stem cell hierarchies in carcinoma-derived cell lines. Cancer research. 2005; 65:8944-8950.
10. Morrison SJ and Kimble J. Asymmetric and symmetric stem-cell divisions in development and cancer. Nature. 2006; 441:1068-1074.
11. Tang DG. Understanding cancer stem cell heterogeneity and plasticity. Cell research. 2012; 22:457-472.
12. Enderling H. Cancer stem cells and tumor dormancy. Advances in experimental medicine and biology. 2013; 734:55-71.
13. Li L and Bhatia R. Stem cell quiescence. Clinical cancer research. 2011; 17:4936-4941.
14. Zeuner A, Francescangeli F, Contavalli P, Zapparelli G, Apuzzo T, Eramo A, Baiocchi M, De Angelis ML, Biffoni M, Sette G, Todaro M, Stassi G and De Maria R. Elimination of quiescent/slow-proliferating cancer stem cells by Bcl-X inhibition in non-small cell lung cancer. Cell Death & Differentiation. 2014; 21:1877-88.
15. Mertins SD. Cancer stem cells: a systems biology view of their role in prognosis and therapy. Anticancer Drugs. 2014; 25:353-367.
16. Zeuner A, Francescangeli F, Contavalli P, Zapparelli G, Apuzzo T, Eramo A, Baiocchi M, De Angelis ML, Biffoni M, Sette G, Todaro M, Stassi G and De Maria R. Elimination of quiescent/slow-proliferating cancer stem cells by Bcl-XL inhibition in non-small cell lung cancer. Cell Death & Differentiation. 2014; 21:1877-1888.
17. Kleffel S and Schatton T. Tumor dormancy and cancer stem cells: two sides of the same coin? Advances in experimental medicine and biology. 2013; 734:145-179.
18. Flesken-Nikitin A, Hwang CI, Cheng CY, Michurina TV, Enikolopov G and Nikitin AY. Ovarian surface epithelium at the junction area contains a cancer-prone stem cell niche. Nature. 2013; 495:241-245.
19. Giancotti FG. Mechanisms governing metastatic dormancy and reactivation. Cell. 2013; 155:750-764.
20. Medema JP and Vermeulen L. Microenvironmental regulation of stem cells in intestinal homeostasis and cancer. Nature. 2011; 474:318-326.
21. Shestopalov IA and Zon LI. Stem cells: The right neighbour. Nature. 2012; 481:453-455.
22. Blagosklonny MV. Target for cancer therapy: proliferating cells or stem cells. Leukemia. 2006; 20:385-391.
23. Blagosklonny MV. Cancer stem cell and cancer stemloids: from biology to therapy. Cancer biology & therapy. 2007; 6:1684-1690.
24. Gupta PB, Fillmore CM, Jiang G, Shapira SD, Tao K, Kuperwasser C and Lander ES. Stochastic state transitions give rise to phenotypic equilibrium in populations of cancer cells. Cell. 2011; 146:633-644.
25. Chaffer CL, Brueckmann I, Scheel C, Kaestli AJ, Wiggins PA, Rodrigues LO, Brooks M, Reinhardt F, Su Y, Polyak K, Arendt LM, Kuperwasser C, Bierie B, et al. Normal and neoplastic nonstem cells can spontaneously convert to a stem-like state. Proceedings of the National Academy of Sciences of the United States of America. 2011; 108:7950-7955.
26. Chaffer CL, Marjanovic ND, Lee T, Bell G, Kleer CG, Reinhardt F, D’Alessio AC, Young RA and Weinberg RA. Poised chromatin at the ZEB1 promoter enables breast cancer cell plasticity and enhances tumorigenicity. Cell. 2013; 154:61-74.
27. Schwitalla S, Fingerle AA, Cammareri P, Nebelsiek T, Goktuna SI, Ziegler PK, Canli O, Heijmans J, Huels DJ, Moreaux G, Rupec RA, Gerhard M, Schmid R, et al. Intestinal tumorigenesis initiated by dedifferentiation and acquisition of stem-cell-like properties. Cell. 2013; 152:25-38.
28. He K, Xu T and Goldkorn A. Cancer cells cyclically lose and regain drug-resistant highly tumorigenic features characteristic of a cancer stem-like phenotype. Molecular cancer therapeutics. 2011; 10:938-948.
29. Magbanua MJ, Carey LA, DeLuca A, Hwang J, Scott JH, Rimawi MF, Mayer EL, Marcom PK, Liu MC, Esteva FJ, Park JW, Rugo HS and Translational Breast Cancer Research C. Circulating tumor cell analysis in metastatic triple-negative breast cancers. Clinical cancer research. 2015; 21:1098-1105.
30. Hsieh JC, Lin HC, Huang CY, Hsu HL, Wu TM, Lee CL, Chen MC, Wang HM and Tseng CP. Prognostic value of circulating tumor cells with podoplanin expression in patients with locally advanced or metastatic head and neck squamous cell carcinoma. Head & neck. 2015; 37:1448-1455.
31. Fu L, Liu F, Fu H, Liu L, Yuan S, Gao Y, Fu Z and Yu J. Circulating tumor cells correlate with recurrence in stage III small-cell lung cancer after systemic chemoradiotherapy and prophylactic cranial irradiation. Japanese journal of clinical oncology. 2014; 44:948-955.
32. Goldkorn A, Ely B, Quinn DI, Tangen CM, Fink LM, Xu T, Twardowski P, Van Veldhuizen PJ, Agarwal N, Carducci MA, Monk JP 3rd, Datar RH, Garzotto M, et al. Circulating tumor cell counts are prognostic of overall survival in SWOG S0421: a phase III trial of docetaxel with or without atrasentan for metastatic castration-resistant prostate cancer. Journal of clinical oncology. 2014; 32:1136-1142.
33. Liao WT, Ye YP, Deng YJ, Bian XW and Ding YQ. Metastatic cancer stem cells: from the concept to therapeutics. American journal of stem cells. 2014; 3:46-62.
34. Mitra A, Mishra L and Li S. EMT, CTCs and CSCs in tumor relapse and drug-resistance. Oncotarget. 2015; 6:10697-10711. doi: 10.18632/oncotarget.4037.
35. Barriere G, Fici P, Gallerani G, Fabbri F, Zoli W and Rigaud M. Circulating tumor cells and epithelial, mesenchymal and stemness markers: characterization of cell subpopulations. Annals of translational medicine. 2014; 2:109.
36. Theodoropoulos PA, Polioudaki H, Agelaki S, Kallergi G, Saridaki Z, Mavroudis D and Georgoulias V. Circulating tumor cells with a putative stem cell phenotype in peripheral blood of patients with breast cancer. Cancer letters. 2010; 288:99-106.
37. Balic M, Lin H, Young L, Hawes D, Giuliano A, McNamara G, Datar RH and Cote RJ. Most early disseminated cancer cells detected in bone marrow of breast cancer patients have a putative breast cancer stem cell phenotype. Clinical cancer research. 2006; 12:5615-5621.
38. Kang Y and Pantel K. Tumor cell dissemination: emerging biological insights from animal models and cancer patients. Cancer cell. 2013; 23:573-581.
39. Bao B, Ahmad A, Azmi AS, Ali S and Sarkar FH. Overview of cancer stem cells (CSCs) and mechanisms of their regulation: implications for cancer therapy. Current protocols in pharmacology / editorial board, SJ Enna. 2013; Chapter 14:Unit 14 25.
40. Aktas B, Tewes M, Fehm T, Hauch S, Kimmig R and Kasimir-Bauer S. Stem cell and epithelial-mesenchymal transition markers are frequently overexpressed in circulating tumor cells of metastatic breast cancer patients. Breast cancer research. 2009; 11:R46.
41. Marusyk A and Polyak K. Tumor heterogeneity: causes and consequences. Biochimica et biophysica acta. 2010; 1805:105-117.
42. Jamal-Hanjani M, Quezada SA, Larkin J and Swanton C. Translational implications of tumor heterogeneity. Clinical cancer research. 2015; 21:1258-1266.
43. Mroz EA, Tward AD, Hammon RJ, Ren Y and Rocco JW. Intra-tumor genetic heterogeneity and mortality in head and neck cancer: analysis of data from the Cancer Genome Atlas. PLoS medicine. 2015; 12:e1001786.
44. Meacham CE and Morrison SJ. Tumour heterogeneity and cancer cell plasticity. Nature. 2013; 501:328-337.
45. Pece S, Tosoni D, Confalonieri S, Mazzarol G, Vecchi M, Ronzoni S, Bernard L, Viale G, Pelicci PG and Di Fiore PP. Biological and molecular heterogeneity of breast cancers correlates with their cancer stem cell content. Cell. 2010; 140:62-73.
46. Pattabiraman DR and Weinberg RA. Tackling the cancer stem cells - what challenges do they pose? Nature reviews Drug discovery. 2014; 13:497-512.
47. Shuang ZY, Wu WC, Xu J, Lin G, Liu YC, Lao XM, Zheng L and Li S. Transforming growth factor-beta1-induced epithelial-mesenchymal transition generates ALDH-positive cells with stem cell properties in cholangiocarcinoma. Cancer letters. 2014; 354:320-328.
48. Bessede E, Staedel C, Acuna Amador LA, Nguyen PH, Chambonnier L, Hatakeyama M, Belleannee G, Megraud F and Varon C. Helicobacter pylori generates cells with cancer stem cell properties via epithelial-mesenchymal transition-like changes. Oncogene. 2014; 33:4123-4131.
49. Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, Campbell LL, Polyak K, Brisken C, et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008; 133:704-715.
50. Kaiser J. The cancer stem cell gamble. Science. 2015; 347:226-229.
51. Chen K, Huang YH and Chen JL. Understanding and targeting cancer stem cells: therapeutic implications and challenges. Acta Pharmacologica Sinica. 2013; 34:732-740.
52. Besancon R, Valsesia-Wittmann S, Puisieux A, Caron de Fromentel C and Maguer-Satta V. Cancer stem cells: the emerging challenge of drug targeting. Current Medicinal Chemistry. 2009; 16:394-416.
53. Plaks V, Kong N and Werb Z. The cancer stem cell niche: how essential is the niche in regulating stemness of tumor cells? Cell stem cell. 2015; 16:225-238.
54. Filatova A, Acker T and Garvalov BK. The cancer stem cell niche(s): the crosstalk between glioma stem cells and their microenvironment. Biochimica et biophysica acta. 2013; 1830:2496-2508.
55. Borovski T, De Sousa EMF, Vermeulen L and Medema JP. Cancer stem cell niche: the place to be. Cancer research. 2011; 71:634-639.
56. Nie D. Cancer stem cell and niche. Frontiers in bioscience. 2010; 2:184-193.
57. Maguire R, Kotronoulas G, Simpson M and Paterson C. A systematic review of the supportive care needs of women living with and beyond cervical cancer. Gynecologic oncology. 2015; 136:478-490.
58. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J and Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015; 65:87-108.
59. Cooke SL, Temple J, Macarthur S, Zahra MA, Tan LT, Crawford RA, Ng CK, Jimenez-Linan M, Sala E and Brenton JD. Intra-tumour genetic heterogeneity and poor chemoradiotherapy response in cervical cancer. British journal of cancer. 2011; 104:361-368.
60. Kidd EA and Grigsby PW. Intratumoral metabolic heterogeneity of cervical cancer. Clinical cancer research. 2008; 14:5236-5241.
61. Huang Z, Wu T, Liu AY and Ouyang G. Differentiation and transdifferentiation potentials of cancer stem cells. Oncotarget. 2015; 6:39550-39563. doi: 10.18632/oncotarget.6098.
62. Di Felice E, Caroli S, Paterlini L, Campari C, Prandi S and Giorgi Rossi P. Cervical cancer epidemiology in foreign women in Northern Italy: role of human papillomavirus prevalence in country of origin. European journal of cancer prevention. 2015; 24:223-230.
63. Herfs M, Yamamoto Y, Laury A, Wang X, Nucci MR, McLaughlin-Drubin ME, Munger K, Feldman S, McKeon FD, Xian W and Crum CP. A discrete population of squamocolumnar junction cells implicated in the pathogenesis of cervical cancer. Proceedings of the National Academy of Sciences of the United States of America. 2012; 109:10516-10521.
64. Herfs M and Crum CP. Cervical cancer: squamocolumnar junction ablation--tying up loose ends? Nature reviews Clinical oncology. 2015; 12:378-380.
65. Herfs M, Parra-Herran C, Howitt BE, Laury AR, Nucci MR, Feldman S, Jimenez CA, McKeon FD, Xian W and Crum CP. Cervical squamocolumnar junction-specific markers define distinct, clinically relevant subsets of low-grade squamous intraepithelial lesions. The American journal of surgical pathology. 2013; 37:1311-1318.
66. Casagrande N, De Paoli M, Celegato M, Borghese C, Mongiat M, Colombatti A and Aldinucci D. Preclinical evaluation of a new liposomal formulation of cisplatin, lipoplatin, to treat cisplatin-resistant cervical cancer. Gynecologic oncology. 2013; 131:744-752.
67. Liu SY and Zheng PS. High aldehyde dehydrogenase activity identifies cancer stem cells in human cervical cancer. Oncotarget. 2013; 4:2462-2475. doi: 10.18632/oncotarget.1578.
68. Lopez J, Poitevin A, Mendoza-Martinez V, Perez-Plasencia C and Garcia-Carranca A. Cancer-initiating cells derived from established cervical cell lines exhibit stem-cell markers and increased radioresistance. BMC cancer. 2012; 12:48.
69. Valent P, Bonnet D, De Maria R, Lapidot T, Copland M, Melo JV, Chomienne C, Ishikawa F, Schuringa JJ, Stassi G, Huntly B, Herrmann H, Soulier J, et al. Cancer stem cell definitions and terminology: the devil is in the details. Nature Reviews Cancer. 2012; 12:767-775.
70. Stacy AE, Jansson PJ and Richardson DR. Molecular pharmacology of ABCG2 and its role in chemoresistance. Molecular pharmacology. 2013; 84:655-669.
71. Zhou S, Schuetz JD, Bunting KD, Colapietro AM, Sampath J, Morris JJ, Lagutina I, Grosveld GC, Osawa M, Nakauchi H and Sorrentino BP. The ABC transporter Bcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant of the side-population phenotype. Nature medicine. 2001; 7:1028-1034.
72. Jonker JW, Freeman J, Bolscher E, Musters S, Alvi AJ, Titley I, Schinkel AH and Dale TC. Contribution of the ABC transporters Bcrp1 and Mdr1a/1b to the side population phenotype in mammary gland and bone marrow of mice. Stem cells. 2005; 23:1059-1065.
73. Wang F, Mi YJ, Chen XG, Wu XP, Liu Z, Chen SP, Liang YJ, Cheng C, To KK and Fu LW. Axitinib targeted cancer stemlike cells to enhance efficacy of chemotherapeutic drugs via inhibiting the drug transport function of ABCG2. Molecular medicine. 2012; 18:887-898.
74. Wang DS, Patel A, Shukla S, Zhang YK, Wang YJ, Kathawala RJ, Robey RW, Zhang L, Yang DH, Talele TT, Bates SE, Ambudkar SV, Xu RH, et al. Icotinib antagonizes ABCG2-mediated multidrug resistance, but not the pemetrexed resistance mediated by thymidylate synthase and ABCG2. Oncotarget. 2014; 5:4529-4542. doi: 10.18632/oncotarget.2102.
75. Ding XW, Wu JH and Jiang CP. ABCG2: a potential marker of stem cells and novel target in stem cell and cancer therapy. Life sciences. 2010; 86:631-637.
76. Hegedus C, Telbisz A, Hegedus T, Sarkadi B and Ozvegy-Laczka C. Lipid regulation of the ABCB1 and ABCG2 multidrug transporters. Advances in cancer research. 2015; 125:97-137.
77. Jia Y, Chen J, Zhu H, Jia ZH and Cui MH. Aberrantly elevated redox sensing factor Nrf2 promotes cancer stem cell survival via enhanced transcriptional regulation of ABCG2 and Bcl-2/Bmi-1 genes. Oncology reports. 2015; 34:2296-2304.
78. Villanueva-Toledo J, Ponciano-Gomez A, Ortiz-Sanchez E and Garrido E. Side populations from cervical-cancer-derived cell lines have stem-cell-like properties. Molecular biology reports. 2014; 41:1993-2004.
79. Douville J, Beaulieu R and Balicki D. ALDH1 as a functional marker of cancer stem and progenitor cells. Stem cells and development. 2009; 18:17-25.
80. Ginestier C, Hur MH, Charafe-Jauffret E, Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG, Liu S, Schott A, Hayes D, Birnbaum D, et sl. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell stem cell. 2007; 1:555-567.
81. Neumeister V, Agarwal S, Bordeaux J, Camp RL and Rimm DL. In situ identification of putative cancer stem cells by multiplexing ALDH1, CD44, and cytokeratin identifies breast cancer patients with poor prognosis. The American journal of pathology. 2010; 176:2131-2138.
82. Charafe-Jauffret E, Ginestier C, Bertucci F, Cabaud O, Wicinski J, Finetti P, Josselin E, Adelaide J, Nguyen TT, Monville F, Jacquemier J, Thomassin-Piana J, Pinna G, et al. ALDH1-positive cancer stem cells predict engraftment of primary breast tumors and are governed by a common stem cell program. Cancer research. 2013; 73:7290-7300.
83. Yao T, Wu Z, Liu Y, Rao Q and Lin Z. Aldehyde dehydrogenase 1 (ALDH1) positivity correlates with poor prognosis in cervical cancer. The Journal of international medical research. 2014; 42:1038-1042.
84. Rao QX, Yao TT, Zhang BZ, Lin RC, Chen ZL, Zhou H, Wang LJ, Lu HW, Chen Q, Di N and Lin ZQ. Expression and functional role of ALDH1 in cervical carcinoma cells. Asian Pacific journal of cancer prevention. 2012; 13:1325-1331.
85. Shmelkov SV, St Clair R, Lyden D and Rafii S. AC133/CD133/Prominin-1. The international journal of biochemistry & cell biology. 2005; 37:715-719.
86. Medema JP. Cancer stem cells: the challenges ahead. Nature cell biology. 2013; 15:338-344.
87. Cheng JX, Liu BL and Zhang X. How powerful is CD133 as a cancer stem cell marker in brain tumors? Cancer treatment reviews. 2009; 35:403-408.
88. Liu TJ, Sun BC, Zhao XL, Zhao XM, Sun T, Gu Q, Yao Z, Dong XY, Zhao N and Liu N. CD133+ cells with cancer stem cell characteristics associates with vasculogenic mimicry in triple-negative breast cancer. Oncogene. 2013; 32:544-553.
89. Sahlberg SH, Spiegelberg D, Glimelius B, Stenerlow B and Nestor M. Evaluation of cancer stem cell markers CD133, CD44, CD24: association with AKT isoforms and radiation resistance in colon cancer cells. PloS one. 2014; 9:e94621.
90. Zhang L, Li H, Ge C, Li M, Zhao FY, Hou HL, Zhu MX, Tian H, Zhang LX, Chen TY, Jiang GP, Xie HY, Cui Y, et al. Inhibitory effects of transcription factor Ikaros on the expression of liver cancer stem cell marker CD133 in hepatocellular carcinoma. Oncotarget. 2014; 5:10621-10635. doi: 10.18632/oncotarget.2524.
91. Chai S, Tong M, Ng KY, Kwan PS, Chan YP, Fung TM, Lee TK, Wong N, Xie D, Yuan YF, Guan XY and Ma S. Regulatory role of miR-142-3p on the functional hepatic cancer stem cell marker CD133. Oncotarget. 2014; 5:5725-5735. doi: 10.18632/oncotarget.2167.
92. Cui F, Wang J, Chen D and Chen YJ. CD133 is a temporary marker of cancer stem cells in small cell lung cancer, but not in non-small cell lung cancer. Oncology reports. 2011; 25:701-708.
93. El-Khattouti A, Selimovic D, Haikel Y, Megahed M, Gomez CR and Hassan M. Identification and analysis of CD133(+) melanoma stem-like cells conferring resistance to taxol: An insight into the mechanisms of their resistance and response. Cancer letters. 2014; 343:123-133.
94. Lai CY, Schwartz BE and Hsu MY. CD133+ melanoma subpopulations contribute to perivascular niche morphogenesis and tumorigenicity through vasculogenic mimicry. Cancer research. 2012; 72:5111-5118.
95. Ji YI, Lee BY, Kang YJ, Jo JO, Lee SH, Kim HY, Kim YO, Lee C, Koh SB, Kim A, Lee JY, Jung MH, Ock MS, et al. Expression patterns of Thymosin beta4 and cancer stem cell marker CD133 in ovarian cancers. Pathology oncology research. 2013; 19:237-245.
96. Kim KH, Kang YJ, Jo JO, Ock MS, Moon SH, Suh DS, Yoon MS, Park ES, Jeong N, Eo WK, Kim HY and Cha HJ. DDX4 (DEAD box polypeptide 4) colocalizes with cancer stem cell marker CD133 in ovarian cancers. Biochemical and biophysical research communications. 2014; 447:315-322.
97. Zhou Q, Chen A, Song H, Tao J, Yang H and Zuo M. Prognostic value of cancer stem cell marker CD133 in ovarian cancer: a meta-analysis. International journal of clinical and experimental medicine. 2015; 8:3080-3088.
98. Qi W, Zhao C, Zhao L, Liu N, Li X, Yu W and Wei L. Sorting and identification of side population cells in the human cervical cancer cell line HeLa. Cancer cell international. 2014; 14:3.
99. Wang K, Zeng J, Luo L, Yang J, Chen J, Li B and Shen K. Identification of a cancer stem cell-like side population in the HeLa human cervical carcinoma cell line. Oncology letters. 2013; 6:1673-1680.
100. Kumazawa S, Kajiyama H, Umezu T, Mizuno M, Suzuki S, Yamamoto E, Mitsui H, Sekiya R, Shibata K and Kikkawa F. Possible association between stem-like hallmark and radioresistance in human cervical carcinoma cells. The journal of obstetrics and gynaecology research. 2014; 40:1389-1398.
101. Yu KR, Yang SR, Jung JW, Kim H, Ko K, Han DW, Park SB, Choi SW, Kang SK, Scholer H and Kang KS. CD49f enhances multipotency and maintains stemness through the direct regulation of OCT4 and SOX2. Stem cells. 2012; 30:876-887.
102. Jeter CR, Yang T, Wang J, Chao HP and Tang DG. Concise Review: NANOG in Cancer Stem Cells and Tumor Development: An Update and Outstanding Questions. Stem cells. 2015; 33:2381-2390.
103. Huntsman HD, Bat T, Cheng H, Cash A, Cheruku PS, Fu JF, Keyvanfar K, Childs RW, Dunbar CE and Larochelle A. Human hematopoietic stem cells from mobilized peripheral blood can be purified based on CD49f integrin expression. Blood. 2015; 126:1631-1633.
104. To K, Fotovati A, Reipas KM, Law JH, Hu K, Wang J, Astanehe A, Davies AH, Lee L, Stratford AL, Raouf A, Johnson P, Berquin IM, et al. Y-box binding protein-1 induces the expression of CD44 and CD49f leading to enhanced self-renewal, mammosphere growth, and drug resistance. Cancer research. 2010; 70:2840-2851.
105. Fukamachi H, Seol HS, Shimada S, Funasaka C, Baba K, Kim JH, Park YS, Kim MJ, Kato K, Inokuchi M, Kawachi H, Yook JH, Eishi Y, et al. CD49f(high) cells retain sphere-forming and tumor-initiating activities in human gastric tumors. PloS one. 2013; 8:e72438.
106. Guo C, Liu H, Zhang BH, Cadaneanu RM, Mayle AM and Garraway IP. Epcam, CD44, and CD49f distinguish sphere-forming human prostate basal cells from a subpopulation with predominant tubule initiation capability. PloS one. 2012; 7:e34219.
107. Haraguchi N, Ishii H, Mimori K, Ohta K, Uemura M, Nishimura J, Hata T, Takemasa I, Mizushima T, Yamamoto H, Doki Y and Mori M. CD49f-positive cell population efficiently enriches colon cancer-initiating cells. International journal of oncology. 2013; 43:425-430.
108. Yamamoto H, Masters JR, Dasgupta P, Chandra A, Popert R, Freeman A and Ahmed A. CD49f is an efficient marker of monolayer- and spheroid colony-forming cells of the benign and malignant human prostate. PloS one. 2012; 7:e46979.
109. Nichols J, Zevnik B, Anastassiadis K, Niwa H, Klewe-Nebenius D, Chambers I, Scholer H and Smith A. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell. 1998; 95:379-391.
110. Boyer LA, Lee TI, Cole MF, Johnstone SE, Levine SS, Zucker JP, Guenther MG, Kumar RM, Murray HL, Jenner RG, Gifford DK, Melton DA, Jaenisch R, et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell. 2005; 122:947-956.
111. Yang Y, Wang Y, Yin C and Li X. Clinical significance of the stem cell gene Oct-4 in cervical cancer. Tumour biology : the journal of the International Society for Oncodevelopmental Biology and Medicine. 2014; 35:5339-5345.
112. Shen L, Huang X, Xie X, Su J, Yuan J and Chen X. High Expression of SOX2 and OCT4 Indicates Radiation Resistance and an Independent Negative Prognosis in Cervical Squamous Cell Carcinoma. The journal of histochemistry and cytochemistry. 2014; 62:499-509.
113. Wang YD, Cai N, Wu XL, Cao HZ, Xie LL and Zheng PS. OCT4 promotes tumorigenesis and inhibits apoptosis of cervical cancer cells by miR-125b/BAK1 pathway. Cell death & disease. 2013; 4:e760.
114. Wang X and Dai J. Concise review: isoforms of OCT4 contribute to the confusing diversity in stem cell biology. Stem cells. 2010; 28:885-893.
115. Rijlaarsdam MA, van Herk HA, Gillis AJ, Stoop H, Jenster G, Martens J, van Leenders GJ, Dinjens W, Hoogland AM, Timmermans M and Looijenga LH. Specific detection of OCT3/4 isoform A/B/B1 expression in solid (germ cell) tumours and cell lines: confirmation of OCT3/4 specificity for germ cell tumours. British journal of cancer. 2011; 105:854-863.
116. Liu L, Wu L, Wei X and Ling J. Induced overexpression of Oct4A in human dental pulp cells enhances pluripotency and multilineage differentiation capability. Stem cells and development. 2015; 24:962-972.
117. Li SW, Wu XL, Dong CL, Xie XY, Wu JF and Zhang X. The differential expression of OCT4 isoforms in cervical carcinoma. PloS one. 2015; 10:e0118033.
118. Weber CE, Kothari AN, Wai PY, Li NY, Driver J, Zapf MA, Franzen CA, Gupta GN, Osipo C, Zlobin A, Syn WK, Zhang J, Kuo PC, et al. Osteopontin mediates an MZF1-TGF-beta1-dependent transformation of mesenchymal stem cells into cancer-associated fibroblasts in breast cancer. Oncogene. 2015; 34:4821-4833.
119. Sakaguchi H, Fujimoto J, Hong BL and Tamaya T. Clinical implications of osteopontin in metastatic lesions of uterine cervical cancers. Cancer letters. 2007; 247:98-102.
120. Wohlleben G, Scherzad A, Guttler A, Vordermark D, Kuger S, Flentje M and Polat B. Influence of hypoxia and irradiation on osteopontin expression in head and neck cancer and glioblastoma cell lines. Radiation oncology. 2015; 10:167.
121. Huang X, Qian Y, Wu H, Xie X, Zhou Q, Wang Y, Kuang W, Shen L, Li K, Su J, Shen L and Chen X. Aberrant expression of osteopontin and E-cadherin indicates radiation resistance and poor prognosis for patients with cervical carcinoma. The journal of histochemistry and cytochemistry. 2015; 63:88-98.
122. Raja R, Kale S, Thorat D, Soundararajan G, Lohite K, Mane A, Karnik S and Kundu GC. Hypoxia-driven osteopontin contributes to breast tumor growth through modulation of HIF1alpha-mediated VEGF-dependent angiogenesis. Oncogene. 2014; 33:2053-2064.
123. Kumar V, Behera R, Lohite K, Karnik S and Kundu GC. p38 kinase is crucial for osteopontin-induced furin expression that supports cervical cancer progression. Cancer research. 2010; 70:10381-10391.
124. Cho H, Hong SW, Oh YJ, Kim MA, Kang ES, Lee JM, Kim SW, Kim SH, Kim JH, Kim YT and Lee K. Clinical significance of osteopontin expression in cervical cancer. Journal of cancer research and clinical oncology. 2008; 134:909-917.
125. Cao L, Fan X, Jing W, Liang Y, Chen R, Liu Y, Zhu M, Jia R, Wang H, Zhang X, Zhang Y, Zhou X, Zhao J, et al. Osteopontin promotes a cancer stem cell-like phenotype in hepatocellular carcinoma cells via an integrin-NF-kappaB-HIF-1alpha pathway. Oncotarget. 2015; 6:6627-6640. doi: 10.18632/oncotarget.3113.
126. Pietras A, Katz AM, Ekstrom EJ, Wee B, Halliday JJ, Pitter KL, Werbeck JL, Amankulor NM, Huse JT and Holland EC. Osteopontin-CD44 signaling in the glioma perivascular niche enhances cancer stem cell phenotypes and promotes aggressive tumor growth. Cell stem cell. 2014; 14:357-369.
127. Adachi K, Suemori H, Yasuda SY, Nakatsuji N and Kawase E. Role of SOX2 in maintaining pluripotency of human embryonic stem cells. Genes to cells. 2010; 15:455-470.
128. Fong H, Hohenstein KA and Donovan PJ. Regulation of self-renewal and pluripotency by Sox2 in human embryonic stem cells. Stem cells. 2008; 26:1931-1938.
129. Ji J and Zheng PS. Expression of Sox2 in human cervical carcinogenesis. Human pathology. 2010; 41:1438-1447.
130. Ji J, Wei X and Wang Y. Embryonic stem cell markers Sox-2 and OCT4 expression and their correlation with WNT signal pathway in cervical squamous cell carcinoma. International journal of clinical and experimental pathology. 2014; 7:2470-2476.
131. Cai C, Tan G, Yu Q, Luan F, Yu L and Wang Y. Expression of SOX2 in cervical intraepithelial neoplasia and cervical cancer and its clinical significance [Article in Chinese]. Nan fang yi ke da xue xue bao. 2013; 33:128-130.
132. Ji J, Ning FR, Liu HJ, Wei X, Zhao J and Wang YL. Effect of Sox2 on proliferation of cervical squamous cancer cell line SiHa [Article in Chinese]. Sichuan da xue xue bao Yi xue ban. 2014; 45:785-788.
133. Canham M, Charsou C, Stewart J, Moncur S, Hoodless L, Bhatia R, Cong D, Cubie H, Busby-Earle C, Williams A, McLoughlin V, Campbell JD, Cuschieri K, et al. Increased cycling cell numbers and stem cell associated proteins as potential biomarkers for high grade human papillomavirus+ve pre-neoplastic cervical disease. PloS one. 2014; 9:e115379.
134. Liu XF, Yang WT, Xu R, Liu JT and Zheng PS. Cervical cancer cells with positive Sox2 expression exhibit the properties of cancer stem cells. PloS one. 2014; 9:e87092.
135. Wicha MS. Targeting self-renewal, an Achilles’ heel of cancer stem cells. Nature medicine. 2014; 20:14-15.
136. Siddique HR and Saleem M. Role of BMI1, a stem cell factor, in cancer recurrence and chemoresistance: preclinical and clinical evidences. Stem cells. 2012; 30:372-378.
137. Ding B, Liu P, Liu W, Sun P and Wang CL. Emerging roles of Kruppel-like factor 4 in cancer and cancer stem cells. Asian Pacific journal of cancer prevention. 2015; 16:3629-3633.
138. Zhang Y, Hao J, Zheng Y, Jing D, Shen Y, Wang J and Zhao Z. Role of Kruppel-like factors in cancer stem cells. Journal of physiology and biochemistry. 2015; 71:155-164.
139. Lin CC, Sharma SB, Farrugia MK, McLaughlin SL, Ice RJ, Loskutov YV, Pugacheva EN, Brundage KM, Chen D and Ruppert JM. Kruppel-like factor 4 signals through microRNA-206 to promote tumor initiation and cell survival. Oncogenesis. 2015; 4:e155.
140. Takaishi S, Okumura T, Tu S, Wang SS, Shibata W, Vigneshwaran R, Gordon SA, Shimada Y and Wang TC. Identification of gastric cancer stem cells using the cell surface marker CD44. Stem cells. 2009; 27:1006-1020.
141. Su YJ, Lai HM, Chang YW, Chen GY and Lee JL. Direct reprogramming of stem cell properties in colon cancer cells by CD44. The EMBO journal. 2011; 30:3186-3199.
142. Wang C, Xie J, Guo J, Manning HC, Gore JC and Guo N. Evaluation of CD44 and CD133 as cancer stem cell markers for colorectal cancer. Oncology reports. 2012; 28:1301-1308.
143. Ohata H, Ishiguro T, Aihara Y, Sato A, Sakai H, Sekine S, Taniguchi H, Akasu T, Fujita S, Nakagama H and Okamoto K. Induction of the stem-like cell regulator CD44 by Rho kinase inhibition contributes to the maintenance of colon cancer-initiating cells. Cancer research. 2012; 72:5101-5110.
144. Joshua B, Kaplan MJ, Doweck I, Pai R, Weissman IL, Prince ME and Ailles LE. Frequency of cells expressing CD44, a head and neck cancer stem cell marker: correlation with tumor aggressiveness. Head & neck. 2012; 34:42-49.
145. Baumann M and Krause M. CD44: a cancer stem cell-related biomarker with predictive potential for radiotherapy. Clinical cancer research. 2010; 16:5091-5093.
146. Hiraga T, Ito S and Nakamura H. Cancer stem-like cell marker CD44 promotes bone metastases by enhancing tumorigenicity, cell motility, and hyaluronan production. Cancer research. 2013; 73:4112-4122.
147. Dall P, Heider KH, Hekele A, von Minckwitz G, Kaufmann M, Ponta H and Herrlich P. Surface protein expression and messenger RNA-splicing analysis of CD44 in uterine cervical cancer and normal cervical epithelium. Cancer research. 1994; 54:3337-3341.
148. Olsson E, Honeth G, Bendahl PO, Saal LH, Gruvberger-Saal S, Ringner M, Vallon-Christersson J, Jonsson G, Holm K, Lovgren K, Ferno M, Grabau D, Borg A, et al. CD44 isoforms are heterogeneously expressed in breast cancer and correlate with tumor subtypes and cancer stem cell markers. BMC cancer. 2011; 11:418.
149. Naor D, Sionov RV and Ish-Shalom D. CD44: structure, function, and association with the malignant process. Advances in cancer research. 1997; 71:241-319.
150. Li L, Hao X, Qin J, Tang W, He F, Smith A, Zhang M, Simeone DM, Qiao XT, Chen ZN, Lawrence TS and Xu L. Antibody against CD44s inhibits pancreatic tumor initiation and postradiation recurrence in mice. Gastroenterology. 2014; 146:1108-1118.
151. Dang H, Steinway SN, Ding W and Rountree CB. Induction of tumor initiation is dependent on CD44s in c-Met(+) hepatocellular carcinoma. BMC cancer. 2015; 15:161.
152. Shimabukuro K, Toyama-Sorimachi N, Ozaki Y, Goi T, Furukawa K, Miyasaka M and Aso T. The expression patterns of standard and variant CD44 molecules in normal uterine cervix and cervical cancer. Gynecologic oncology. 1997; 64:26-34.
153. Martinho O, Goncalves A, Moreira MA, Ribeiro LF, Queiroz GS, Schmitt FC, Reis RM and Longatto-Filho A. KIT activation in uterine cervix adenosquamous carcinomas by KIT/SCF autocrine/paracrine stimulation loops. Gynecologic oncology. 2008; 111:350-355.
154. Conic I, Stanojevic Z, Jankovic Velickovic L, Stojnev S, Ristic Petrovic A, Krstic M, Stanojevic M, Bogdanovic D and Stefanovic V. Epithelial ovarian cancer with CD117 phenotype is highly aggressive and resistant to chemotherapy. The journal of obstetrics and gynaecology research. 2015; 41:1630-1637.
155. Chen D, Zhang Y, Wang J, Chen J, Yang C, Cai K, Wang X, Shi F and Dou J. MicroRNA-200c overexpression inhibits tumorigenicity and metastasis of CD117+CD44+ ovarian cancer stem cells by regulating epithelial-mesenchymal transition. Journal of ovarian research. 2013; 6:50.
156. Chen J, Wang J, Chen D, Yang J, Yang C, Zhang Y, Zhang H and Dou J. Evaluation of characteristics of CD44+CD117+ ovarian cancer stem cells in three dimensional basement membrane extract scaffold versus two dimensional monocultures. BMC cell biology. 2013; 14:7.
157. Zhang X, Kyo S, Nakamura M, Mizumoto Y, Maida Y, Bono Y, Takakura M and Fujiwara H. Imatinib sensitizes endometrial cancer cells to cisplatin by targeting CD117-positive growth-competent cells. Cancer Letters. 2014; 345:106-114.
158. Adhikari AS, Agarwal N, Wood BM, Porretta C, Ruiz B, Pochampally RR and Iwakuma T. CD117 and Stro-1 identify osteosarcoma tumor-initiating cells associated with metastasis and drug resistance. Cancer Research. 2010; 70:4602-4612.
159. Rocha-Zavaleta L, Huitron C, Caceres-Cortes JR, Alvarado-Moreno JA, Valle-Mendiola A, Soto-Cruz I, Weiss-Steider B and Rangel-Corona R. Interleukin-2 (IL-2) receptor-betagamma signalling is activated by c-Kit in the absence of IL-2, or by exogenous IL-2 via JAK3/STAT5 in human papillomavirus-associated cervical cancer. Cellular signalling. 2004; 16:1239-1247.
160. Han CP, Chen CK, Lin CK, Wang PH and Chiang H. Unusual c-KIT (+) squamous cell carcinoma of uterine cervix showing remarkable platelet-derived growth factor receptor, alpha subunit expression, but no activating mutation. Histopathology. 2011; 59:788-790.
161. Chao WR, Lin WL, Chen CK, Han LM, Lin JC and Han CP. Unusual c-KIT+ squamous cell carcinoma of the uterine cervix showing paradoxical hypermethylation of the c-KIT proto-oncogene. European journal of obstetrics, gynecology, and reproductive biology. 2015; 184:130-131.
162. Gagliardi A, Mullin NP, Ying Tan Z, Colby D, Kousa AI, Halbritter F, Weiss JT, Felker A, Bezstarosti K, Favaro R, Demmers J, Nicolis SK, Tomlinson SR, et al. A direct physical interaction between Nanog and Sox2 regulates embryonic stem cell self-renewal. The EMBO journal. 2013; 32:2231-2247.
163. Pan G and Thomson JA. Nanog and transcriptional networks in embryonic stem cell pluripotency. Cell research. 2007; 17:42-49.
164. Wang D, Lu P, Zhang H, Luo M, Zhang X, Wei X, Gao J, Zhao Z and Liu C. Oct-4 and Nanog promote the epithelial-mesenchymal transition of breast cancer stem cells and are associated with poor prognosis in breast cancer patients. Oncotarget. 2014; 5:10803-10815. doi: 10.18632/oncotarget.2506.
165. Iv Santaliz-Ruiz LE, Xie X, Old M, Teknos TN and Pan Q. Emerging role of nanog in tumorigenesis and cancer stem cells. International journal of cancer. 2014; 135:2741-2748.
166. Schreiber L, Raanan C and Amsterdam A. CD24 and Nanog identify stem cells signature of ovarian epithelium and cysts that may develop to ovarian cancer. Acta histochemica. 2014; 116:399-406.
167. Di J, Duiveman-de Boer T, Zusterzeel PL, Figdor CG, Massuger LF and Torensma R. The stem cell markers Oct4A, Nanog and c-Myc are expressed in ascites cells and tumor tissue of ovarian cancer patients. Cellular oncology. 2013; 36:363-374.
168. Amsterdam A, Raanan C, Schreiber L, Freyhan O, Fabrikant Y, Melzer E and Givol D. Differential localization of LGR5 and Nanog in clusters of colon cancer stem cells. Acta histochemica. 2013; 115:320-329.
169. Ibrahim EE, Babaei-Jadidi R, Saadeddin A, Spencer-Dene B, Hossaini S, Abuzinadah M, Li N, Fadhil W, Ilyas M, Bonnet D and Nateri AS. Embryonic NANOG activity defines colorectal cancer stem cells and modulates through AP1- and TCF-dependent mechanisms. Stem cells. 2012; 30:2076-2087.
170. Shan J, Shen J, Liu L, Xia F, Xu C, Duan G, Xu Y, Ma Q, Yang Z, Zhang Q, Ma L, Liu J, Xu S, et al. Nanog regulates self-renewal of cancer stem cells through the insulin-like growth factor pathway in human hepatocellular carcinoma. Hepatology. 2012; 56:1004-1014.
171. Noh KH, Lee YH, Jeon JH, Kang TH, Mao CP, Wu TC and Kim TW. Cancer vaccination drives Nanog-dependent evolution of tumor cells toward an immune-resistant and stem-like phenotype. Cancer research. 2012; 72:1717-1727.
172. Chiou SH, Yu CC, Huang CY, Lin SC, Liu CJ, Tsai TH, Chou SH, Chien CS, Ku HH and Lo JF. Positive correlations of Oct-4 and Nanog in oral cancer stem-like cells and high-grade oral squamous cell carcinoma. Clinical cancer research. 2008; 14:4085-4095.
173. Zhang J, Wang X, Li M, Han J, Chen B, Wang B and Dai J. NANOGP8 is a retrogene expressed in cancers. The FEBS journal. 2006; 273:1723-1730.
174. Ye F, Zhou C, Cheng Q, Shen J and Chen H. Stem-cell-abundant proteins Nanog, Nucleostemin and Musashi1 are highly expressed in malignant cervical epithelial cells. BMC cancer. 2008; 8:108.
175. Gu TT, Liu SY and Zheng PS. Cytoplasmic NANOG-positive stromal cells promote human cervical cancer progression. The American journal of pathology. 2012; 181:652-661.
176. Korkaya H, Paulson A, Charafe-Jauffret E, Ginestier C, Brown M, Dutcher J, Clouthier SG and Wicha MS. Regulation of mammary stem/progenitor cells by PTEN/Akt/beta-catenin signaling. PLoS Biology. 2009; 7:e1000121.
177. Dubrovska A, Kim S, Salamone RJ, Walker JR, Maira SM, Garcia-Echeverria C, Schultz PG and Reddy VA. The role of PTEN/Akt/PI3K signaling in the maintenance and viability of prostate cancer stem-like cell populations. Proceedings of the National Academy of Sciences of the United States of America. 2009; 106:268-273.
178. Alkatout I, Kabelitz D, Kalthoff H and Tiwari S. Prowling wolves in sheep’s clothing: the search for tumor stem cells. Biological Chemistry. 2008; 389:799-811.
179. Dean M, Fojo T and Bates S. Tumour stem cells and drug resistance. Nature reviews Cancer. 2005; 5:275-284.
180. Schatton T, Frank NY and Frank MH. Identification and targeting of cancer stem cells. BioEssays : news and reviews in molecular, cellular and developmental biology. 2009; 31:1038-1049.
181. Ho MM, Ng AV, Lam S and Hung JY. Side population in human lung cancer cell lines and tumors is enriched with stem-like cancer cells. Cancer Research. 2007; 67:4827-4833.
182. Wang J, Guo LP, Chen LZ, Zeng YX and Lu SH. Identification of cancer stem cell-like side population cells in human nasopharyngeal carcinoma cell line. Cancer Research. 2007; 67:3716-3724.
183. Singh S, Trevino J, Bora-Singhal N, Coppola D, Haura E, Altiok S and Chellappan SP. EGFR/Src/Akt signaling modulates Sox2 expression and self-renewal of stem-like side-population cells in non-small cell lung cancer. Molecular cancer. 2012; 11:73.
184. Shi Y, Fu X, Hua Y, Han Y, Lu Y and Wang J. The side population in human lung cancer cell line NCI-H460 is enriched in stem-like cancer cells. PLoS One. 2012; 7:e33358.
185. Boesch M, Zeimet AG, Reimer D, Schmidt S, Gastl G, Parson W, Spoeck F, Hatina J, Wolf D and Sopper S. The side population of ovarian cancer cells defines a heterogeneous compartment exhibiting stem cell characteristics. Oncotarget. 2014; 5:7027-7039. doi: 10.18632/oncotarget.2053.
186. Golebiewska A, Brons NH, Bjerkvig R and Niclou SP. Critical appraisal of the side population assay in stem cell and cancer stem cell research. Cell Stem Cell. 2011; 8:136-147.
187. Zheng D, Liao S, Zhu G, Zhou Y and Li G. [Side population cells and progress in cancer stem cell research]. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2014; 39:525-531.
188. Broadley KW, Hunn MK, Farrand KJ, Price KM, Grasso C, Miller RJ, Hermans IF and McConnell MJ. Side population is not necessary or sufficient for a cancer stem cell phenotype in glioblastoma multiforme. Stem Cells. 2011; 29:452-461.
189. Lou D, Wang B, Wang Y and Cao Y. Selective killing of cancer stem cells by a novel dual-targeting strategy. Medical hypotheses. 2012; 79:430-432.
190. Kolev VN, Wright QG, Vidal CM, Ring JE, Shapiro IM, Ricono J, Weaver DT, Padval MV, Pachter JA and Xu Q. PI3K/mTOR dual inhibitor VS-5584 preferentially targets cancer stem cells. Cancer research. 2015; 75:446-455.
191. Srivastava M MC, Roda J, Jie HB, Axelrod F, Xie MH, Yun R, Mayes E, Bentley T, Cancilla B, Tam R, Tang T, Kapoun A, Lewick Ji, et al. Dual targeting of Delta-like ligand 4 (DLL4) and programmed death 1(PD1) inhibits tumor growth and generates enhanced long-term immunological memory [abstract]. In: Proceedings of the 106th Annual Meeting of the American Association for Cancer Research. 2015; 75:Abstract nr 255.
192. Rao W, Wang H, Han J, Zhao S, Dumbleton J, Agarwal P, Zhang W, Zhao G, Yu J, Zynger DL, Lu X and He X. Chitosan-Decorated Doxorubicin-Encapsulated Nanoparticle Targets and Eliminates Tumor Reinitiating Cancer Stem-like Cells. ACS nano. 2015; 9:5725-5740.
193. Burke AR, Singh RN, Carroll DL, Torti FM and Torti SV. Targeting Cancer Stem Cells with Nanoparticle-Enabled Therapies. Journal of molecular biomarkers & diagnosis. 2012; Suppl 8.
194. Bale SS, Kwon SJ, Shah DA, Banerjee A, Dordick JS and Kane RS. Nanoparticle-mediated cytoplasmic delivery of proteins to target cellular machinery. ACS nano. 2010; 4:1493-1500.
195. El-Sadik AO, El-Ansary A and Sabry SM. Nanoparticle-labeled stem cells: a novel therapeutic vehicle. Clinical pharmacology : advances and applications. 2010; 2:9-16.
196. Rotherham M and El Haj AJ. Remote activation of the Wnt/beta-catenin signalling pathway using functionalised magnetic particles. PloS one. 2015; 10:e0121761.
197. Yang H, Li Y, Li T, Xu M, Chen Y, Wu C, Dang X and Liu Y. Multifunctional core/shell nanoparticles cross-linked polyetherimide-folic acid as efficient Notch-1 siRNA carrier for targeted killing of breast cancer. Scientific reports. 2014; 4:7072.
198. Dou QQ, Rengaramchandran A, Selvan ST, Paulmurugan R and Zhang Y. Core-shell upconversion nanoparticle - semiconductor heterostructures for photodynamic therapy. Scientific reports. 2015; 5:8252.
199. Kim KS, Park W and Na K. Gadolinium-chelate nanoparticle entrapped human mesenchymal stem cell via photochemical internalization for cancer diagnosis. Biomaterials. 2015; 36:90-97.
200. Wang L, Su W, Liu Z, Zhou M, Chen S, Chen Y, Lu D, Liu Y, Fan Y, Zheng Y, Han Z, Kong D, Wu JC, et al. CD44 antibody-targeted liposomal nanoparticles for molecular imaging and therapy of hepatocellular carcinoma. Biomaterials. 2012; 33:5107-5114.
201. Hong IS, Jang GB, Lee HY and Nam JS. Targeting cancer stem cells by using the nanoparticles. International journal of nanomedicine. 2015; 10:251-260.
202. Ambasta RK, Sharma A and Kumar P. Nanoparticle mediated targeting of VEGFR and cancer stem cells for cancer therapy. Vascular cell. 2011; 3:26.
203. Dhar S, Reddy EM, Prabhune A, Pokharkar V, Shiras A and Prasad BL. Cytotoxicity of sophorolipid-gellan gum-gold nanoparticle conjugates and their doxorubicin loaded derivatives towards human glioma and human glioma stem cell lines. Nanoscale. 2011; 3:575-580.
204. Burke AR, Singh RN, Carroll DL, Wood JC, D’Agostino RB Jr, Ajayan PM, Torti FM and Torti SV. The resistance of breast cancer stem cells to conventional hyperthermia and their sensitivity to nanoparticle-mediated photothermal therapy. Biomaterials. 2012; 33:2961-2970.
205. Satapathy SR, Siddharth S, Das D, Nayak A and Kundu CN. Enhancement of Cytotoxicity and Inhibition of Angiogenesis in Oral Cancer Stem Cells by a Hybrid Nanoparticle of Bioactive Quinacrine and Silver: Implication of Base Excision Repair Cascade. Molecular pharmaceutics. 2015;12:4011-25. doi: 10.1021/acs.molpharmaceut.5b00461.
206. Deonarain MP, Kousparou CA and Epenetos AA. Antibodies targeting cancer stem cells: a new paradigm in immunotherapy? mAbs. 2009; 1:12-25.